Вырожденная селекция групп в головном мозгу
На этом фоне мы теперь можем подробнее охарактеризовать систему вырожденной селекции групп, которая, как мы полагаем, развилась для осуществления высших функций мозга. Прежде всего надо сказать, что мы ограничим наше обсуждение областями коры, таламусом и лимбической системой; функции других частей нервной оси можно будет рассмотреть впоследствии.
Первый важный вопрос состоит в следующем: что распознает ЦНС? То есть, каков элементарный субстрат высших функций мозга? Мы примем здесь, что выше того уровня, где внешний сигнал взаимодействует с сенсорными рецепторами, которые могут действовать линейным или нелинейным образом (Mountcastle, 1957, 1967), субстрат для распознавания состоит из пространственно-временных последовательностей спайков и конфигураций градуальных потенциалов (и сопутствующих им химических превращений), происходящих в группах нейронов. Под распознаванием я понимаю избирательную и характерную дискриминативную реакцию одной или нескольких групп нейронов на такие конфигурации, вызываемые или наличные в других группах, к которым они имеют доступ. Реакция таких распознающих групп выражается в изменении характера их последующей активности и в стабильных или метастабильных синаптических изменениях. Теоретическое описание возможных внутренних свойств нейронных групп, на которых могло бы основываться распознавание между группами, дают Уилсон и Коуэн (Wilson, Cowan, 1973). Важно подчеркнуть, что сделанные допущения не исключают распознавания состояния отдельного нейрона в иерархии. Но в общем предполагается, что сигналы, распознаваемые клеточными группами, генерируются другими группами. Некоторое подтверждение этого положения дают работы Хьюбела и Визела (Hubel, Wiesel, 1974) по обнаружению признаков в стриарной коре и связанных с ней областях.
Хотя о существовании клеточных групп в других областях имеются лишь скудные данные, может быть, на фоне ранее проведенного анализа (Bullock, 1961) для большей ясности полезно шире рассмотреть понятия кле-
Дж. Эделтен
точной группы, ассортимента и распознавания. Группой клеток называется набор смежных нейронов, внутренние связи в котором определены событиями, происходившими в онтогенезе и во время развития. Связи внутри группы не случайные, а определенные. Каждая такая группа может обладать дивергирующими или конвергирующими внешними связями, идущими к другим таким группам и от них; эти связи тоже нейроанатомически определены и не случайны. Согласно такому представлению, хотя одиночный нейрон редко может служить «группой», случайно составленные сети исключены (Bullock, 1961). В соответствии с проводимыми Баллоком (там же) различиями группа могла бы составлять метастабильную петлю обратной связи с множественными входами, состоящую из сети взаимодействующих нейронов с определенными свойствами. Но мы предполагаем наличие у групп нескольких дополнительных свойств, которые не предусматривались прежними теоретиками (Bullock, 1961; Grosser, Griisser-Cornehls, 1976).
Одно из самых важных таких свойств заключается в том, что группы образуют наборы, т. е. совокупности клеток с разпыми внутренними, но сходными наружными связями. Такие наборы становятся специализированными в процессе онтогенеза и являются вырожденными в отношении распознавания. Каждая группа в наборе может действовать как единица распознавания и выполнять одну или несколько функций: кодирование, декодирование, идентификацию одной из линий ее внешних связей, распределение сигналов во времени, определение их силы или скорости их развития или продолжительности — все это могут быть свойства сигналов, приходящих от других клеточных групп. Разумеется, в большинстве отдельных случаев точный нейронный код в настоящее время неизвестен.
Если внешние связи группы дивергируют и ведут ко многим другим группам, то выходная активность группы может мобилизовать другие группы более или менее эффективно в зависимости от многих разнообразных факторов, таких, как структура сигнала, сила сигнала и локальное торможение. Как в случае суда присяжных, эти группы могут ответить или не ответить характерным об-
Селекция групп и фазная повторная сигнализация 83
разом на входную активность первой группы. Если внешние связи конвергируют на группе, то возможно множество разных реакций, в том числе облегчение, временные различия или различные структуры реакций, причем все они зависят от расположения и свойств внешних с.вязей. Таким образом, в группе могут быть разные ворота для входного притока, и приток к разным комбинациям этих ворот может влиять или не влиять на реакцию группы. Благодаря структуре своих внутренних и внешних связей, нейроны группы обладают многообразными способами взаимодействия с внешними сигналами. Но отдельная группа не способна давать множество разных реакций, напротив, она располагает ограниченным набором характерных пространственно-временных форм импульсных реакций, а также характерным для нее набором связей с другими группами.
Среди различных видов изменчивости, представляемых таким набором связей, самой важной для обеспечения функций высшего порядка является возможность пластических изменений внутренних связей в синапсах — комми-тирование («commitment»), что ведет к стабилизации определенной формы выхода. Об этом подробнее будет сказано ниже. А здесь для нас главное то, что изменения такого рода, происходящие главным образом на внутренних синапсах, должны давать преимущество одним внутренним связям над другими.
Теперь мы можем рассмотреть иерархию реакций, которая на своих поздних стадиях нелинейна из-за наличия петель обратных и прямых связей с присущими им изменениями временной последовательности и времени реакции. Пренебрежем ненадолго этой нелинейностью и рассмотрим иерархию
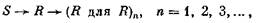
где S — это преобразованный сенсорный входной приток из окружающей среды (как только один пример входа), R — группы клеток коры, которые могут действовать как «распознаватели» этого входа (например, группы сложных нейронов в стриарной коре), а (Л для R) означает группы нейронов в ассоциативной, или височной, или лобной, или префронтальной коре, которые действуют как
Дж. Эделтен
«распознаватели для распознавателей». Согласно этой иерархии, сигналы от нейронных групп/? (например, колонки) могут быть распознаны группами в (R для R). Но следует подчеркнуть, что на этом высшем уровне распознавания группы нейронов — кандидаты в (R для R) — образуют лишь вырожденную подгруппу всех клеточных групп (R для R) (рис. 4, А). Иными словами, (R для R)
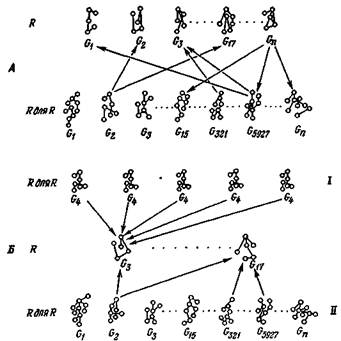
Йю. 4. Взаимодействие вырожденных групп в двух ассортиментах R и (R для R). Стрелки, соединяющие группы, означают распознавание, но не обязательно предполагают передачу информации только в одном направлении. А. Вырожденность; специально покапаны двусторонние отношения по принципу связи одной группы с несколькими. Б. Различие между избыточностью (/) и вырожденностью (//) в (R для R). Увеличение числа избыточных элементов может лишь повысить эффективность; так, из-за своего строения и функции G4 —полностью избыточная группа ассортимента (Я для R) — никогда не сможет распознать G17 в R.
Селекция групп и фазная повторная сигнализация 85
содержит не одну, а несколько групп, которые способны распознать определенную группу в R; эта реакция основана на возможности дивергенции и на множественном вырожденном представительстве, которые вместе обеспечивают адекватное распознавание групп R в ассортименте (R для R). Заметим также, что стрелки на этой схеме означают необязательно поток информации в одном направлении, а всего лишь исходную последовательность событий распознавания. Действительно, схему не следует рассматривать как строго или исключительно иерархическую. Совершенно очевидно, что так же важна параллельная организация, поскольку она делает возможным выход на действие (двигательную реакцию, нейрогуморальную реакцию) с любого уровня. Схема предполагает также, что общие свойства входа могут быть распознаны на раннем этапе в последовательности S—>-/?—>(R для R).
Здесь мы можем подвести итог, сказав в несколько общей форме, что выше уровня сенсорного преобразования и сенсорной обработки ЦНС распознает собственные операции избирательно, нелинейно, вырожденно. Полезно задать вопрос, может ли такое вырожденное распознавание успешно осуществляться изоморфными, или избыточными (в строгом смысле слова), группами клеток в (R для R). При условии что пороговое распознавание не требует абсолютной специфичности, а лишь различения между классами сходных структур сигналов, ответ на этот вопрос по причинам, приведенным в предыдущем разделе, будет отрицательным (рис. 4, Б). Напротив, требуется значительное число разных, но изофункциональных групп клеток, каждая из которых может реагировать на данный класс структур в R. Это означает, что одна и та же структура сигнала может быть распознана несколькими способами и что при последовательных предъявлениях одной и той же структуры реагировать будут разные комбинации клеточных групп (R для R). Более того, нет нужды принимать a priori, что такие группы обязательно состоят из одинакового числа или одинаковых типов нейронов.
Такую картину нейронных групп, отобранных из вырожденного ассортимента, надо отличать от фиксированной структуры заранее запрограммированной реакции,
Дж. Эделмен
которая может возникнуть в отдельном ганглии, а также от простой избыточности нейронов в таких субсистемах, как ганглии или ядерные клеточные массы головного мозга. Хотя такие организации в головном мозгу несомненно имеются, они не могут генерировать такого рода реакции, какие формируются вырожденным ассортиментом.
До сих нор я лишь мельком касался различий между начальным устойчивым состоянием и последующими реактивными состояниями подгрупп клеточных групп в таком наборе. Согласно постулированной теории, описанный ассортимент можно считать первичным ассортиментом, возникшим в процессе онтогенеза и раннего развития как следствие дифференцировки, которая привела к образованию синаптических связей — как локальных, так и в длинных проводящих путях. Однако, как уже указывалось, если только в этом наборе не возникла какая-либо форма стабильного усиления (или торможения) реакций клеточных групп этого набора, изменение его свойств под воздействием входных сигналов будет невозможно. Дело ограничится обратимой флуктуацией его избирательной реакции с различными интервалами расслабления. Здесь мы должпы предположить, что селекция некоторых подгрупп изменяет вероятность того, что эти подгруппы снова будут выбраны при повторном предъявлении стимула подобной структуры. Как было сказано выше, предполагается, что это происходит вследствие синаптических изменений некоторых или всех клеток группы, из-за чего внутренняя или внешняя нейронная сеть функционально меняется. Следует ожидать, что это происходит главным образом во внутренней сети группы при сопутствующем изменении, например, ее передаточных функций. Вероятность селекции может понизиться (торможение) или возрасти (возбуждение, облегчение). В обоих случаях достаточно частое повторение входного сигнала за определенное время, по предположению, изменяет вероятность отбора в будущем некоторых ранее выбранных подгрупп предпочтительно перед их соседями — процесса, создающего вторичный ассортимент. Таким образом, вторичный ассортимент — это совокупность разных групп нейронов высшего порядка, у которых внутренняя или внешняя синаптическая функция изменена отбором (се-
Селекция групп и фазная повторная сигнализация 87
лекцией) и коммитированием в процессе активности. Кроме того, повторение входных сигналов необязательно ограничивается внешними стимулами — оно может относиться к повторным входам, идущим от самого мозга.
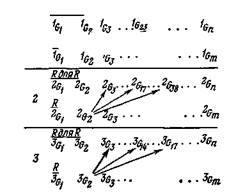
Можно представить себе нелинейную реакцию, построенную по принципу связи многих элементов с одним, зависимую от времени и вырожденную (рис. 5); здесь верхними индексами обозначены последовательные структурированные состояния, а нижними индексами — разные клеточные группы в наборе. Повышенная вероятность положительной или отрицательной селекции представлена на рисунке жирной стрелкой.
Изменение этой вероятности может быть связано с порогом распознавания. Благодаря тому что группы клеток с большей частотой повторения реакций лучше «подходят», соответствуют, входным сигналам, они стабилизируются более эффективно. Это может происходить во внутренних элементах, но скорее всего посредством избавления от торможения. В то же время тормозный сиг-
Rd/inR 1 я /^^
Дж. Эделмен
нал, который подавил активность некоторых распознающих клеточных групп, слабее отвечающих на первоначальный вход, должен эффективно обострить специфичность суммарного ответа, а также усилить его стабилизацию. Таким образом, селекция клеточных групп может происходить путем фильтрации — сначала путем стимуляции групп, которые реагируют более или менее хорошо, а затем путем торможения (или конкурентного исключения) выбранных групп с недостаточной реакцией относительно некоторого порога (рис. 6).
Теперь мы можем заняться вопросом, касающимся общих свойств такой системы, и затем рассмотреть данные в пользу существования клеточных групп — кандидатов на обладание такими свойствами. Очевидно, что изменения вероятностей селекции представляют собой феномен памяти; на рис. 6 не выделен ассоциативный характер этой памяти (Languet-Higgins, Willshow, Berneman, 1970), проистекающий из свойств вырожденной селекции.
fy Ц2 2ез"'_________• • • 3?я
