Образование РНК-затравки на о.ц. ДНК-матрице
Лекция 3
1.Образование РНК-затравки на о.ц. ДНК-матрице а) с помощью РНК полимеразы б) с помощью праймазыPapovaviridae
Polioma Vacuolating virises (SV 40 (Simian Vacuolating Virus)) Размер генома примерно одинаков: вирусы папилломы – 8 т.п.н., полиомы – 5 т.п.н. Детально мы рассмотрим вирусы полиомы…SV 40.
Геном представлен двуцепочечной кольцевой ДНК. В геноме закодирован белок – большой Т-Ag (80 кДа). Он единственный вирусный белок участвующий в репликации ДНК. Белок полифункционален:
1.способен взаимодействовать с рядом TF (переход клетки в S-фазу)
2.сам является TF (синтез вирусных белков)
3.узнает ori (т.е. отностся к группе белков OBP-Ori Binding Proteins)
4.способен к олигомеризации. В присутствии АТФ и ori образует кольцевой гексамер.
5.обладает хеликазной активностью (3'-5')
Примечание: может неспецифически взаимодействовать с о.ц.ДНК.
|
| |||||||||||||||||||||
 | |||||||||||||||||||||
 |  |  |  | ||||||||||||||||||
| |||||||||||||||||||||
 | |||||||||||||||||||||
 | |||||||||||||||||||||
 | |||||||||||||||||||||
| |||||||||||||||||||||
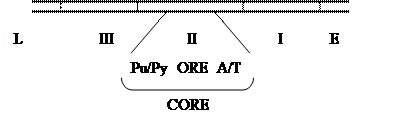
 ori состоит из трех участков. В этих находятся пентануклеотидные последовательности GAGGC ( ), узнаваемые T-Ag. В III участке находятся повторы 21, 22 п.н.
ori состоит из трех участков. В этих находятся пентануклеотидные последовательности GAGGC ( ), узнаваемые T-Ag. В III участке находятся повторы 21, 22 п.н.
Во II втором участке находится ORE (Ori Recognition Element), с которым связывается T-Ag, он входит в состав CORE (Core Ori Recognition Element) длиной 64 п.н., и фланкирован c одной стороны А/Т богатым участком, с другой - Pu/Py последовательностью. Общая длина ori со всеми прилегающими участками – 170 п.н.
Сила взаимодействия T-Agс ORE зависит от степени его фосфорилирования.
Инициация репликации SV 40.
| ||||||
 | ||||||
 | ||||||
 | ||||||
Далее T-Ag проявляет хеликазную активность (3'-5') и расплетает область Ori. На о.ц. ДНК тут же садятся олигомеры RPA (Replicating protein A, аналог SSB E.coli). Затем с расплетенным участком связывается ДНКpolα/праймаза. В комплексе устанавливается физическое взаимодействие T-Ag с RPA и ДНКpolα. ДНКpolα синтезирует РНК-ДНК затравку: короткий олигорибонуклеотид (10 нт) и короткий олигодезоксирибонуклеотид (~20 нт).
T-Ag специфически узнает клеточные белки RPA и ДНКpolα только приматов. Этим и ограничен круг хозяев SV40.
Примечание: внешние факторы, определяющие хозяева вирусов – белки оболочки, в данном случае мы встречаемся с внутренними факторами.
В комплекс входят также клеточная топоизомераза I, устраняющая топологические проблемы (также взаимодействует с Т-Ag) и протеинфосфатаза, регулирующая степень фосфорилированности T-Ag.
Примечание: ДНК SV40 упакована в нуклеосомы (около 20 нуклеосом), т.е. представляет собой минихромосому. Одна из нуклеосом позиционирована на ori и делает его недоступным для взаимодействия с T-Ag. Освобождают ori клеточные комплексы ремоделирования.
Инициация репликации у вирусов полиомы.
Все в принципе то же самое отличие в том, что вместо пентануклеотидных повторов в ori находятся гексануклеотидные и их положение немного отличается.
Элонгация репликации SV40.
Примечание: при репликации ДНК одна из синтезируемых цепей является "лидирующей", она синтезируется непрерывно, другая цепь называется… Умозрительное объяснение этого факта – модель "трубы". В результате работы комплекса, включающего T-Ag и полимеразы, образуется репликативный " глазок ". По мере…Терминация репликации ДНК SV40.
В итоге образуется…Фаг Р4.
Линейная д.ц. ДНК длиной около 12 т.п.н. с липкими…Фаг λ.
А/Т rich Структура ori. …I2 R3 I1 R2 R1
 |
I1 , I2 - узнаются IHF (изгибает ДНК на 270˚С)
R1 , R2 , R3 - узнаются Nu1
Терминаза в виде димера присоединяется к CosB и на расстоянии 12 п.н. вносит разрыв в CosN. С этого участка начинается упаковка ДНК. CosQ нужен для правильного процессирования противоположного конца ДНК, чтобы упаковался точно один эквивалент генома. Поскольку для начала упаковки ДНК и завершения этого процесса используются сигналы одного и того же элемента, то должна быть регуляция специфичности внесения разрыва терминазой. Возможно, процесс регулируется путем взаимодействия белков капсида с терминазой.
Фаг Р22 (лямбдоидный фаг).
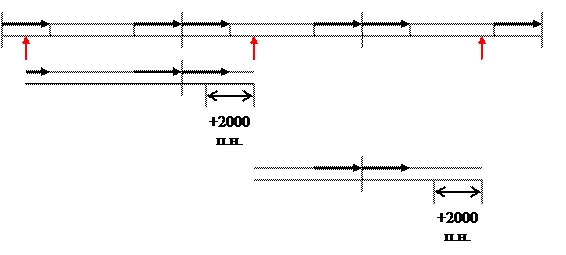
Геном представлен линейной двуцепочечной ДНК с тупыми концами. На концах прямые повторы длиной более 2000 п.н. Последовательность концевых повторов не фиксирована из-за кольцевых пермутаций. Причем пермутации полные, а у фага λ разброс в пределах 20%.

Как и у фага λ ori состоит из 4 итеронов, с прлегающей к ним А/Т-богатой областью. Ori находится внутри гена 18, гомологичного гену О, который кодирует аналог белка О фага λ. Рядом находится ген 12, кодирующий хеликазу.
На поздней стадии репликации трудно обнаружить кольцевые молекулы, но конкатемеры образуются. Может быть, конкатемеры образуются не по схеме Кернса.
Имеется белок угнетающий активность клеточной системы RecBCD. Упаковка ДНК начинается с сайта Pac разрыв вносится собственной терминазой. Работой упаковочной системы можно объяснить кольцевые пермутации. Имеется несколько Pac сайтов, разбросанных по геному. Pac сайт первого эквивалента генома выбирается случайно. Второй разрыв вносится после полного заполнения головки (fullhead упаковка). Помещается больше, чем эквивалент генома на длину прямого повтора (2000 п.н.). Упаковка следующего эквивалента генома начинается с конца первого (упаковка процессивна).