рефераты конспекты курсовые дипломные лекции шпоры

Реферат Курсовая Конспект
Условия, достаточные для сознательного восприятия
Условия, достаточные для сознательного восприятия - Лекция, раздел Медицина, Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга ВНашем Изложении Мы До Сих Пор Не Касались Всех Условий, Дос...
Внашем изложении мы до сих пор не касались всех условий, достаточных для состояния сознания и близкой к этому проблемы качества сенсорных модальностей. В некотором отношении вопрос качества субъективен и недоступен научному тестированию. Проверка возможна
Дж. Эделмен
в лучшем случае посредством отчетов, но прямое сравнение сенсорных качеств, одинаково определяемых двумя людьми, еще не может служить проверкой их качественного сходства или различия. Это представляется тем более вероятным, если принять во внимание большое разнообразие состояний двух сознательных субъектов. Но все же рассмотрение некоторых аспектов сенсорного качества в свете селективной теории функции мозга может принести определенную пользу.
Для начала стоит указать, что, поскольку сенсорные модальности передаются сигналами по «меченым линиям», в принципе не существует проблемы идентификации разных модальностей. Однако их распознавание группами R и (R для R) происходит посредством ряда иерархически упорядоченных абстрактных преобразований, возникающих в результате активности этих групп. Согласно селективной теории, одновременности входов Si, £2, ..., Sn достаточно, чтобы повысить возможность ассоциации высшего порядка между соответствующими результатами их преобразований; при этом необязательно, чтобы эти S были причинно связаны.
Если иметь это в виду, то весьма вероятно, что условия, достаточные для формирования сознания, возникают из исторического процесса в каждом индивидууме, причем все более абстрактные операции помещаются во вторичном ассортименте. Следует, однако, учитывать еще одну возможность, а именно что ранние ассоциации с теми областями мозга, которые связаны с эффективными состояниями, являются решающими для распределения в памяти ряда комбинаций (patterns) реакций, которые часто используются в жизни в дальнейшем. Эти распределенные «примитивы» могут состоять из модально связанных форм моторпых и сенсорных реакций, которые первоначально осуществлялись, например, через посредство областей гипоталамуса и медиального пучка переднего мозга. Такие комбинации реакций могут приводить к происходящим при участии химических медиаторов изменениям многих соматических реакций. Хотя мы не в состоянии рассмотреть их «качество» в научных терминах, так как доступ к этим реакциям возможен только косвенный (посредством наблюдения за поведением) или вербальный (может
Селекция групп и фазная повторная сигналигация 111
быть, в такое время, когда вербальное общение еще не развилось), позднее «качества», описываемые вербально, могут быть привязаны к своим реакциям посредством абстракции через (R для R)-группы. Способность различать модальности может играть в этом процессе главную роль, поскольку ассоциативные кросс-корреляции модальностей создаются благодаря одновременности разных входов S. Так или иначе выражение сенсорного качества представляет собой процесс в высшей степени абстрактный, на что указывали Миллер и Джонсон-Лэрд (Miller, Johnson-Laird, 1976). Принимая во внимание данные, полученные во многих работах (Piagel, 1950, 1954; Miller, Johnson-Laird, 1976), это также означает, что осознание качества является исторически развившимся процессом.
На рис. 13 показано последующее обращение к разным иерархиям (R для R) путем кросс-корреляции S и взаимодействия с «примитивами» в ассортименте. Из-за сложности таких взаимодействий этот рисунок и допущения, на которых он основан, надо принимать как весьма предварительную схему.
Как ясно из всего сказанного, если настаивать на том, что условия, достаточные для состояния сознания, должны включать объяснение качества в том смысле, что оно дается индивидууму в опыте путем непосредственного ознакомления, то в таком случае построение удовлетворительной научной теории невозможно. Поэтому предлагаемая теория не дает такого определения сознания, которое удовлетворяло бы требованиям достаточности подобного рода, как их предписывают философы. Но все же можно сказать, что в предлагаемой модели исключены некоторые условия, которые ранее вызывали трудности:
1. Необходимость в «думающем гомункулусе» отпадает, если постулируются связи между фазной обработкой повторных входов и абстрактной многомерной памятью, благодаря которым «я» и модель мира описываются в понятиях прошлого сенсорного и моторного опыта. В своих наиболее изощренных формах такие модели, вероятно, потребуют наличия элементов, способных к речи (Miller, Johnson-Laird, 1967), но речь, возможно, не всегда обязательна для их проявления.
2. Требование бесконечного регресса или иерархии
Дж. Эделмен
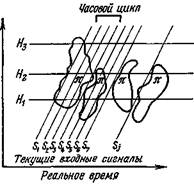
Рис. 13. Попытка показать, как последовательные циклы фазного повторного входа с ассоциативным считыванием могут направить вызов на иерархические уровни нейронов (R для R) возрастающих степеней абстрактного кодирования. Для «примитивов» П, состоящих из распределенных заложенных в памяти следов прежнего опыта, ассоциированного в разной степени с различными иерархическими уровнями Hi—Д3, характерна высокая вероятность взаимодействия с текущим входом и считывания нейронами (R для R). Три произвольно выбранных иерархических уровня хранения состоят из групп нейронов (R для R), которые содержат все более абстрактную информацию, и поэтому они представляют собой разнообразные рутинные операции, накопленные в опыте. Более высокие уровни содержат более абстрактные стабильные представительства, которые не в такой мере зависят от непосредственного изменения во времени. Например, Ht может представлять мгновенное распознавание признака и образа; Я2 — ассоциативные и перцептивные рутинные схемы; Н3 — внутренние модели или «теории» личности, возможно базирующиеся на лингвистических рутинных операциях. Ассоциация текащего входа S, на уровнях Hi и Н2, ведущая к корреляции модальности, может быть прочитана группами, которые ассоциированы с Я3. Временные отношения, модальность и аффективные связи могут привести к разным двигательным реакциям, зависящим от доступа к «примитивам». Часовой цикл включает несколько одновременных, но не синхронизированных S/.
устраняется вырожденностью и ассоциативными свойствами групп клеток. Далее, оно снимается фазным повторным характером сигнализации, который допускает «повторные старты» без утраты возможности обращения к долговременной памяти. Дивергентные (вырожденные) и конвергентные (абстрагирующие) свойства (R дляЛ)-ан-самблей позволяют думать, что с гнездами абстракций
Селекция групп и фазная повторная сигнализация 113
высшего порядка может иметь дело последовательный процесс. Кроме того, благодаря ассоциативным свойствам вырожденной селекции групп ослабляется потребность в программе или «программисте». В этом смысле аналогия между высшей нервной деятельностью и компьютерами несостоятельна, хотя на высших уровнях такая система способна выполнять рутинные операции (Miller, Johnson-Laird, 1976).
3. К входным сигналам не предъявляются требования причинности: достаточно одновременности при условии соблюдения временных констант церебральных, ретикулярных и лимбических реакций.
Согласно этой точке зрения, сознательное восприятие требует временных процессов, которые и параллельны, и последовательны. Происходит непрерывное челночное движение кросс-коррелированных сигналов, которые по отдельным фазам обращаются к исторически сложившейся памяти. А в операциональном смысле если сконструировать машину с такими свойствами, то, согласно этой точке зрения, она будет описывать состояния сознания (или обнаруживать их при тестировании). Но выявляемое «путем ознакомления» качество, присущее ощущению или восприятию, не будет прямо доступно и его нельзя будет операционально определить, если только машина не будет соединена с нервной системой человека, что еще менее вероятно, чем само конструирование машины. О таких качествах машина могла бы сказать: «Если бы они не были такими, то были бы этакими». Главное условие адекватною функционирования состоит в том, чтобы, получив сигналы меченых модальностей, машина не путала их.
Некоторые следствия и сравнения с другими точками зрения
Нужно указать на некоторые общие следствия, вытекающие из этой модели. Во-первых, нет нужды постулировать, что на молекулы головного мозга непосредственно влияют процессы сознания, как об этом говорит Сперри (Sperry, 1969, 1970), выдвигая представление о модифицированном ментализме. Помимо того, что эти допущения сформулированы в термодинамически неясных или сомни-
Дж. Эделмен
тельных терминах, они не нужны как и акцентирование особенных свойств «мозга» и холистские объяснения, лишенные механических деталей. Предлагаемая схема не требует также построения дуалистических или плюралистических моделей (Eccles, 1974; Popper, 1974) и не опирается на определенные состояния с психофизическим параллелизмом. В отличие от этого занятая нами позиция согласуется в основном с позицией так называемого материализма центрального состояния (Campbell, 1970) или с более сложными вариантами гипотезы «идентичности» (Feigl, 1967). Однако в качестве научной теории наши построения предлагают для объяснения высших функций мозга особый комплекс механизмов, и победа или поражение этой теории зависит от возможности их проверки, а не от ее философского альянса.
Было бы ошибкой сделать из приведенного описания вывод о том, что система с вырожденной селекцией групп и повторным входом сигналов работает по принципу часового механизма. Незначительная флуктуация внешнего сигнала порождает в активирующих системах или в (R для R) очень много изофункциопальпых, но неидентичных (R для R)-реакций на генерирующие состояния, из которых могут возникнуть новые селективные реакции. Селекция из клеточных групп, участвующих в этих состояниях, может идти без того, чтобы нужно было «говорить молекулам, что им делать». На этом уровне описания наиболее существенными элементами процесса сознания являются клеточные группы, их связи и генерируемый ими разнообразный репертуар. Поскольку существует огромный набор возможностей для сравнения состояний таких групп с последующими состояниями, особенно при небольших отклонениях внешних сигналов, нет необходимости обращаться к детерминированным «механическим» последовательностям реакций. Эю представляется особенно верным, если принять во внимание, что два эквивалентных состояния могут представлять совершенно разные подгруппы (R для R)-ассортимента. Важнее то, что перекомбинация таких состояний породит почти безграничное число возможных ассоциаций.
При наличии долговременной памяти в (R для R) и способности изменять степень обработки S и активирую-
Селекция групп и фазная повторная сигнализация 115
щих сигналов, нетрудно представить себе, как можно было бы модулировать такую систему для планирования и программирования, для ограничения сигналов проприо-цептивными входами или для акцентирования взаимодействий (R для R) при измененных состояниях внимания (Виноградова, 1975). В этой связи нужно снова подчеркнуть, что в любое время результат процесса, протекающего в системе вырожденных групп с фазным повторным входом, вероятностно предопределен. Так, конкурентная селекция между или среди групп первичного и вторичного ассортимента будет определяться: 1) процессом мобилизации, т. е. тем, какую достаточно «подходящую» группу или группы клеток первыми встретил данный сигнал; 2) наличием тормозных процессов, которые устраняют реакции групп с меньшей степенью распознавания, и 3) степенью коммитирования и эффективностью распознавания предварительно выбранной группы во вторичном ассортименте. Для зрелой системы это не исключает возможности внутренней генерации S. И действительно, замена «внешнего» S (или конкуренция с ним) внутренне генерируемым S важна для центрального планирования и программирования. Но я опасаюсь, что для этого нужны значительный предварительный опыт селекции и обработка высшего порядка, которая связана в своих самых изощренных формах с приобретением речи (Miller, Johnson-Laird, 1976). Это требует a fortiori тонких различий между «я» и «не-я» (Piaget, 1950, 1954).
В любом случае главное состоит в том, что сознание предполагает избирательные фазные взаимодействия как с памятью, так и с внешними входными сигналами и что она полностью зависит от прошлого и настоящего опыта. Такая система обладает свободой («свободой воли» или «свободой действия»)—свобода заложена в селективной грамматике нейронных групп, — но эта свобода не безгранична, она лимитирована различными запретами, которые налагаются свойствами групп нейронов.
– Конец работы –
Эта тема принадлежит разделу:
Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга
На сайте allrefs.net читайте: " Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Условия, достаточные для сознательного восприятия
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.022 сек.
Новости и инфо для студентов