рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Физика
- /
- Структура биологических мембран
Реферат Курсовая Конспект
Структура биологических мембран
Структура биологических мембран - раздел Физика, Биофизика Первая Модель Строения Биологических Мембран Была Предложена В 1902 Г. Было З...
Первая модель строения биологических мембран была предложена в 1902 г. Было замечено, что через мембраны лучше всего проникают вещества, хорошо растворимые в липидах, и на основании этого было сделано предположение, что биологические мембраны состоят из тонкого слоя фосфолипидов. На самом деле, на поверхности раздела полярной и неполярной среды (например, воды и воздуха) молекулы фосфолипидов образуют мономолекулярный (одномолекулярный) слой. Их полярные "головы" погружены в полярную среду, а неполярные "хвосты" ориентированы в сторону неполярной среды. Поэтому и можно было предположить, что биологические мембраны построены из монослоя липидов.
В 1925 г. Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. Гортер и Грендел экстрагировали липиды из гемолизированных эритроцитов ацетоном, затем выпаривали раствор на поверхности воды и измеряли площадь образовавшейся мономолекулярной пленки липидов. На основании результатов этих исследований была высказана идея, что липиды в мембране располагаются в виде бимолекулярного слоя.
Биологическую мембрану можно рассматривать как электрический конденсатор, в котором пластинами являются электролиты наружного и внутреннего растворов (внеклеточного и цитоплазмы) с погруженными в них головами липидных молекул. Проводники разделены диэлектрическим слоем, образованным неполярной частью липидных молекул - двойным слоем их хвостов. Липиды - диэлектрики с диэлектрической проницаемостью e » 2.
Емкость плоского конденсатора:
C = eeoS/d
где электрическая постоянная e0= 8,85 ´ 1012 Ф/м, d - расстояние между пластинами конденсатора, S - площадь пластины.
Удельная емкость (на единицу площади):
Cуд = eeo/d.
Отсюда можно найти расстояние между пластинами конденсатора, соответствующее в нашем случае толщине липидной части мембраны:
d = eeo/Cуд = 8,85´10-12´2/0,5´10-2=3,5 нм.
Это как раз соответствует по порядку величины толщине неполярной части бимолекулярного слоя липидов, сложенных определенным образом.
Однако мембрана - это не только липидный бислой, она состоит и из белковых молекул. При измерении поверхностного натяжения клеточных мембран обнаружено, что измеренные значения коэффициента поверхностного натяжения значительно ближе к коэффициенту поверхностного натяжения на границе раздела белок-вода (около 10-4 Н/м), нежели на границе раздела липид-вода (около 10-2 Н/м). Даниелли и Девсоном предложили в 1935 г. так называемую бутербродную модель строения биологических мембран, которая с некоторыми несущественными изменениями продержалась в мембранологии в течение почти 40 лет. Согласно этой модели мембрана - трехслойная. Она образована двумя расположенными по краям слоями белковых молекул с липидным бислоем посередине; образуется нечто вроде бутерброда: липиды, наподобие масла, между двумя "ломтями" белка.
Однако по мере накопления экспериментальных данных пришлось, в конце концов, отказаться от бутербродной модели строения биологических мембран. Огромную роль в развитии представлений о строении биологических мембран сыграло все большее проникновение в биологию физических методов исследования.
Большую информацию о структуре мембран, о взаимном расположении атомов мембранных молекул дает рентгеноструктурный анализ, основанный на дифракции коротковолновых рентгеновских лучей на атомарных структурах. Исследования дифракции рентгеновских лучей на мембране подтвердили относительно упорядоченное расположение липидных молекул в мембране - двойной молекулярный слой с более или менее параллельно расположенными жирнокислыми хвостами, дали возможность точно определить расстояние между полярной головой липидной молекулы и метильной группой в конце углеводородной цепи.
Наибольшие успехи в раскрытии особенностей строения биологических мембран были достигнуты в электронно-микроскопических исследованиях. Как известно, световой микроскоп не позволяет рассмотреть детали объекта, меньшие примерно половины длины световой волны (около 200 нм). В световом микроскопе можно разглядеть отдельные клетки, однако, он совершенно непригоден для изучения биологических мембран, толщина которых в 20 раз меньше предела разрешения светового микроскопа. Разрешающая способность микроскопа ограничена явлением дифракции. Поэтому, чем меньше длина волны по сравнению с деталями исследуемого объекта, тем меньше искажения. Предел разрешения пропорционален длине волны.
В электронном микроскопе вместо светового пучка на исследуемый объект направляется пучок электронов, разогнанных до больших скоростей. Известно, что электронам с высокими скоростями тоже присущи волновые свойства, в том числе явление дифракции. Однако при достаточно больших скоростях, согласно формуле де Бройля, длина волны мала и соответственно мал предел разрешения. Так, если электроны ускоряются электрическим полем с напряжением 105 В, их скорость достигает 106 м/с, длина волны уменьшается и предел разрешения составляет порядка 0,1 нм, что позволяет рассмотреть отдельные детали строения биологических мембран. В электронном микроскопе достигается увеличение в сотни тысяч раз, что дало возможность исследовать строение клетки, клеточных органелл и биологических мембран.
Недостатком электронной микроскопии является деформация живого объекта в процессе исследования. Перед началом электронномикроскопических исследований клетка проходит через многие стадии предварительной обработки: обезвоживание, закрепление, ультратонкий срез, обработка препаратов веществами, хорошо рассеивающими электроны (например, золотом, серебром, осмием, марганцем и т.п.). При этом изучаемый объект значительно изменяется. При помощи электронной микроскопии удалось получить изображение биологических мембран, на снимках видно трехслойное строение мембраны. Новая информация о строении мембраны была получена с помощью метода "замораживание-скол-травление". По этому методу клетку охлаждают до очень низкой температуры в жидком азоте. Охлаждение проводится с очень большой скоростью (около 1000 градусов в секунду). При этом вода, содержащаяся в препарате, переходит в твердое аморфное состояние. Затем клетки раскалываются специальным ножом и помещаются в вакуум. Замерзшая вода быстро возгоняется, освобождая поверхность скола (этот процесс и называют травлением). После травления получают реплику (отпечаток со сколотой поверхности) и фотографируют в электронном микроскопе. Замороженные мембраны могут при раскалывании расщепляться в разных направлениях, в том числе и вдоль границы двух липидных монослоев, и поэтому можно видеть их внутреннее строение. Было обнаружено, что имеются белковые молекулы, погруженные в липидный бислой и даже прошивающие его насквозь. Это привело к существенному изменению представлений о строении мембраны.
Современное представление о структуре мембраны. Совокупность результатов, полученных физическими и химическими методами исследования, дала возможность предложить новую жидкостно-мозаичную модель строения биологических мембран (Сингер и Никольсон, 1972 г.). Согласно Сингеру и Никольсону, структурную основу биологической мембраны образует двойной слой фосфолипидов, инкрустированный белками.
Различают поверхностные (или периферические) и интегральные белки.

Рис. 3.3 - Структура плазматической мембраны (Кольман Я, Рём К.-Г., 2000)
Липиды находятся при физиологических условиях в жидком агрегатном состоянии. Это позволяет сравнить мембрану с фосфолипидным морем, по которому плавают белковые "айсберги". Одним из подтверждений жидкостно-мозаичной модели является и тот факт, что, как установил химический анализ, в разных мембранах соотношение между содержанием белков и фосфолипидов сильно варьирует: в миелиновой мембране белков в 2,5 раза меньше, чем липидов, а в эритроцитах, напротив, белков в 2,5 раза больше, чем липидов. При этом, согласно современной модели, соотношение количества белков и липидов во всех мембранах должно быть примерно одинаково. Тот факт, что не вся поверхность биологической мембраны покрыта белками, показал и метод ядерного магнитного резонанса.

Рис.3.4 - Мембранные липиды (Кольман Я, Рём К.-Г., 2000)
Кроме фосфолипидов и белков, в биологических мембранах содержатся и другие химические соединения. В мембранах животных клеток много холестерина (в сравнимом количестве с фосфолипидами и белками). Есть в мембранах и другие вещества, например гликолипиды, гликопротеиды.
Жидкостно-мозаичная модель строения мембраны в настоящее время общепринята. Однако, как всякая модель, она дает довольно упрощенную картину строения мембраны. В частности, обнаружено, что белковые "айсберги" не всегда свободно плавают в липидном море, а могут быть "заякорены" на внутренние (цитоплазматические) структуры клетки. К таким структурам относятся микрофиламенты и микротрубочки. Микротрубочки - полые цилиндры диаметром около 300 нм из особого белка (тубулина) играют важную роль в функционировании клетки.
Выяснилось также, что не все липиды в мембране расположены по принципу бислоя. Физические методы исследования показали, что липидная фаза мембран содержит также участки, где липидные молекулы не образуют двойной слой.
Изучением сложного химического состава мембран, мембраных белков и других веществ занимается биохимия. Основная область приложения биофизики - структурная основа мембны, а именно двойной слой фосфолипидных молекул.
Молекула фосфолипида лецитина содержит полярную голову (производную фосфорной кислоты) и длинный неполярный хвост (остатки жирных кислот). В голове фосфолипидной молекулы лецитин имеются две заряженные группы, расположенные на некотором расстоянии друг от друга. Два разноименных заряды, равные по абсолютной величине, образуют электрический диполь.

Рис.3.5 - Мембранные белки (Кольман Я, Рём К.-Г., 2000)
В мембранах содержатся разные фосфолипиды. Например, в мембране эритроцитов их около 20 видов. Варьирует химическая формула полярной головы молекулы. У некоторых фосфолипидов головы кроме двух зарядов противоположного знака, создающих дипольный момент, но оставляющих молекулу в целом нейтральной, несут один отрицательный некомпенсированный заряд, вследствие чего молекула оказывается заряженной отрицательно. Углеводородные хвосты фосфолипидной молекулы содержат приблизительно 20 атомов углерода, в хвосте может быть 1-4 двойных ненасыщенных связей два хвоста.
Полярные головы молекул фосфолипидов - гидрофильны, а их неполярные хвосты - гидрофобны. В смеси фосфолипидов с водой термодинамически выгодно, чтобы полярные головы были погружены в состоящую из полярных молекул воду, а их неполярные хвосты были бы расположены подальше от воды. Такое расположение амфифильных (имеющих и гидрофильную, и гидрофобную части) молекул соответствует наименьшему значению энергии Гиббса по сравнению с другими возможными расположениями молекул.
Очень существенным является то обстоятельство, что молекулы фосфолипидов имеют два хвоста. Такая молекула в пространстве имеет форму, близкую к цилиндру. Из молекул фосфолипидов в водной среде происходит самосборка бислойной мембраны. Присутствие молекул с одним хвостом (лизолецитин), имеющих в пространстве форму, близкую к конусу, разрушает клеточные мембраны. Фосфолипидные молекулы, лишенные одного из хвостов, образуют поры в бислойной мембране, т.е. нарушается барьерная функция мембран.
3.3 Динамика мембран. Подвижность фосфолипидных молекул в мембранах
Режим функционирования мембраны сильно зависит от: микровязкости липидного бислоя и подвижности фосфолипидных молекул в мембране, фазового состояния мембранных липидов. Отклонения биофизических характеристик липидного бислоя от нормы связано с разного рода патологиями. Важную роль в физиологии клетки играют фазовые перекоды в биологических мембранах.
Липидная фаза биологических мембран при физиологических условиях (температуре, давлении, химическом составе окружающей среды) находится в жидком агрегатном состоянии. Это доказано методами флюоресцентного анализа (с использованием флюоресцентных зондов и меток), электоонного парамагнитного резонанса (ЭПР), с использованием «спиновых зондов и меток, и ядерного магнитного резонанса (ЯМР).
В нормальном состоянии мембрана не флюоресцирует. Чтобы провести исследования мембраны флюоресцентным методом, надо вводить в мембрану молекулы или молекулярные группы, способные к флюоресценции. В качестве флюоресцентных зондов используются: ДМХ - диметиламинохалкон; МБА - 3-метоксибензантрон; АНС - 1-анилин-нафталин-сульфонат и др.
Флюоресцентный анализдает возможность исследовать подвижность фосфолипидных молекул в мембране, оценить вязкость липидной фазы мембраны (так называемую микровязкость мембран). Микровязкость мембраны можно оценить по изменениям спектров флюоресценции, а также по степени поляризации Р флюоресцентного излучения при освещении мембраны поляризованным светом. Связь степени поляризации Р и микровязкости мембраны h выражается формулой Перрена и Яблонского:
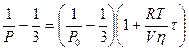 ,
,
где P0 - степень поляризации света на неподвижных молекулах, R = 8,31 Дж / (К ´ моль) - универсальная газовая постоянная, Т [К] - температура, V - молярный объем флюоресцирующих молекул, i время жизни возбужденного состояния.
Наиболее полные сведения об агрегатном состоянии липидных бислоев дают методы радиоспектроскопии ЭПР и ЯМР.
Как и в случае ЭПР, спектры ЯМР тем шире, чем больше вязкость и меньше молекулярная подвижность исследуемого объекта. Флюоресцентные, ЭПР- и ЯМР-исследования показали, что подвижность фосфолипидных молекул в мембране сравнительно велика, а вязкость мала. В нормальных физиологических условиях липидная часть мембраны находится в жидком агрегатном состоянии. Вязкость липидной мембраны сравнима с вязкостью подсолнечного масла:
h » (30-100)мПа´с
(для сравнения: вязкость воды при 20 °С составляет 1 мПа ´ с). Изменение микровязкости липидного окружения мембранных белков-ферментов резко сказывается на их функционировании. Некоторые экспериментальные данные свидетельствуют о том, что канцерогенез связан со снижением вязкости липидной фазы мембраны, а при старении вязкость, напротив, увеличивается. Разрабатываются диагностические методы, основанные на измерении микровязкости мембран с помощью спин-зондов.
Микровязкость мембраны у концов липидных хвостов меньше, чем около полярных голов, высокая подвижность липидных молекул обусловливает латеральную (боковую) диффузию. Латеральная диффузия – это хаотическое тепловое перемещение молекул липидов и белков в плоскости мембраны. При латеральной диффузии рядом расположенные молекулы липидов скачком меняются местами и вследствие таких последовательных перескоков из одного места в другое молекула перемещается вдоль поверхности мембраны. Среднее квадратичное перемещение Sв молекул при диффузии за время t можно оценить по формуле Эйнштейна:
Sкв = 2´(Dt)1/2
Зная skb, можно найти значение коэффициента латеральной диффузии D.
Оказалось, что среднее квадратичное перемещение за секунду фосфолипидной молекулы по поверхности мембраны эритроцита составило около 5 мкм, что сравнимо с размерами клеток. Таким образом, за секунду молекула может обежать всю поверхность небольшой клетки. Обнаруженное среднее квадратичное перемещение белковых молекул составило около 0,2 мкм за секунду.
Рассчитанные по формуле Эйнштейна коэффициенты латеральной диффузии для липидов Dл»6´10-12 м2 / с, для белков Dб»10-14 м2 /c.
Частота перескоков (число перескоков в секунду) молекулы с одного места на другое вследствие латеральной диффузии может быть найдена по формуле:
n = 2´31/2´D/f
где f - площадь, занимаемая одной молекулой на мембране.
Для молекул фосфолипидов Dлип= 6 10-12м2/с, f »7´10-19м2. Для этих значений частота перескоков n = 3 ´ 107 с-1. Каждая молекула, таким образом, в среднем претерпевает десятки миллионов перестановок в плоскости мембраны за секунду, то есть характерное время одного перескока i= 10-7 – 10-8 с.
Флип-флоп - это диффузия молекул мембранных фосфолипидов поперек мембраны.
Скорость перескоков молекул с одной поверхности мембраны на другую (флип-флоп) определена методом спиновых меток в опытах на модельных липидных мембранах – липосомах. Перескоки молекул с одной поверхности бис-лоя на другую совершаются значительно медленнее, чем перескоки при латеральной диффузии. Среднее время, через которое фосфолипидная молекула совершает флип-флоп (Т ~ 1 час), в десятки миллиардов раз больше среднего времени, характерного для перескока молекулы из одного места в соседнее в плоскости мембраны. Сочетание быстрой диффузии молекул вдоль мембраны и очень медленной диффузии поперек мембраны имеет большое значение для функционирования мембран, а именно для матричной функции мембраны. Благодаря затрудненному переходу поперек мембраны поддерживается упорядоченность в молекулярной структуре мембраны, ее анизотропия, асимметрия (относительно плоскости мембраны) расположения липидных и белковых молекул, определенная ориентация белков-ферментов поперек мембраны. Это имеет большое значение, например, для направленного переноса веществ через мембрану.
– Конец работы –
Эта тема принадлежит разделу:
Биофизика
На сайте allrefs.net читайте: Биофизика. Биофизика...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Структура биологических мембран
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.024 сек.
Новости и инфо для студентов