рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Медицина
- /
- Лекция 3. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ГЕНЕТИКИ
Реферат Курсовая Конспект
Лекция 3. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ГЕНЕТИКИ
Лекция 3. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ГЕНЕТИКИ - раздел Медицина, Генетика. Курс лекций для специальности "БИОЭКОЛОГИЯ" Вопросы Лекции: 1. Общая Характеристика Нуклеиновых Кислот...
Вопросы лекции:
1. Общая характеристика нуклеиновых кислот
2. Дезоксирибонуклеиновая кислота (ДНК)
3. Репликация, репарация и рекомбинация ДНК
Общая характеристика нуклеиновых кислот.Нуклеиновые кислоты были открыты швейцарским биологом Фридрихом Мишером в 1868г. при попытке раскрыть химическую природу клеточного ядра. В дальнейшем нуклеиновые кислоты были выявлены во всех клетках человека, животных и растений, а также в микроорганизмах и вирусах.
Существуют два типа нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). Они выполняют следующие функции:
1. Обеспечивают хранение и передачу наследственной информации, непосредственно принимая участие в механизмах реализации этой информации путем программирования синтеза всех клеточных белков.
2. Структурные компоненты нуклеиновых кислот выполняют функции кофакторов (коэнзим А, уридиндифосфатглюкоза, НАД, ФАД и др.).
3. Нуклеиновые кислоты являются макроэргическими соединениями, аккумулируют, переносят и трансформируют энергию (АТФ, АДФ, АМФ) и тем самым принимают участие в биоэнергетике всех живых организмов.
4. Являются предшественниками вторичных посредников (мессенджеров) – циклических мононуклеотидов (цАМФ и цГМФ), которые образуются при гидролизе ДНК и участвуют в передаче внутриклеточных сигналов, например, при действии гормонов.
5. ДНК – это нуклеиновая кислота, мономерами которой являются дезоксирибонуклеотиды. Она является химической основой генов, в которых сконцентрирована наследственная информация организма.
Дезоксирибонуклеиновая кислота (ДНК).В 1924 г. Фёльген доказал, что ДНК является обязательным компонентом хромосом. В 1944 г. О. Эвери, Маклеод и Маккарти установили, что ДНК играет решающую роль в хранении, передаче и реализации наследственной информации. В 1953 г. Фрэнсис Крик и Джеймс Уотсон установили, что молекула ДНК представляет собой двойную спираль полинуклеотидных цепей, закрученных вокруг одной оси.
В зависимости от степени гидратации молекулы ДНК могут существовать в А-, В- и крайне редко Z-формах.
В природе преобладает В-ДНК, которая образуется при содержании в клетке более 40% воды. Это правозакрученная спираль, у которой расстояние между витками (шаг спирали) равно 3,4 нм. На этом участке укладывается 10 нуклеотидных остатков, размер одного нуклеотида составляет 0,34 нм. Диаметр биспиральной молекулы равен 1,8 нм.
А-ДНК представляет собой правозакрученную двойную спираль, у которой меньше шаг спирали и больше диаметр. Z-ДНК – левозакрученная спираль с большим шагом и меньшим диаметром.
Длина ДНК измеряется числом нуклеотидных пар (нп). У мухи дрозофилы она равна 105 нп, у человека – 2900 нп, у дрожжей – 13,5 миллионов нп.
Абсолютное количество нуклеотидов, входящих в состав ДНК колеблется в широких диапазонах, но при этом количество пуриновых и пиримидиновых оснований всегда комплементарно или соответственно.
Репликация ДНК.Репликация или самоудвоение ДНК – один из важнейших биологических процессов, обеспечивающих воспроизведение генетической информации. В результате репликации одной молекулы ДНК образуется две новые молекулы, которые являются точной копией исходной молекулы или матрицы.
Реакции, в которых одна молекула гетерополимера служит матрицей для синтеза другой молекулы гетерополимера с комплементарной структурой, называются реакциями матричного типа. Если в ходе реакции образуются молекулы того же вещества, которое служит матрицей, то реакция называется автокаталитической. Если же в ходе реакции на матрице одного вещества образуются молекулы другого вещества, то такая реакция называется гетерокаталитической. Таким образом, репликация ДНК является автокаталитической реакцией матричного синтеза.
К реакциям матричного типа относятся, в первую очередь, репликация ДНК (синтез ДНК на матрице ДНК), транскрипция ДНК (синтез РНК на матрице ДНК) и трансляция РНК (синтез балка на матрице РНК). Однако существуют и другие реакции матричного типа, например, синтез РНК на матрице РНК и синтез ДНК на матрице РНК. Эти типы реакций наблюдаются при заражении клетки определенными вирусами. Теоретически различают три механизма матричного синтеза ДНК:
1. консервативный, который предусматривает образование дочерней молекулы ДНК на родительской молекуле ДНК без разделения последней;
2. полуконсервативный, который характеризуется расхождением цепей родительской ДНК и синтезом на них двух дочерних цепей, содержащих одну родительскую и одну новую цепь;
3. дисперсный, который связан с расщеплением родительской ДНК в нескольких местах и синтезом на ней новых цепей ДНК, при этом каждая дочерняя цепь ДНК состоит из коротких соединенных между собой участков родительских и новых цепей ДНК.
В 1957 г. М. Мезельсон и Ф. Сталь экспериментально подтвердили существование метода полуконсервативного матричного синтеза ДНК в клетках кишечной палочки.
Все матричные процессы протекают в три этапа: инициация (начало), элонгация (продолжение) и терминация (окончание).
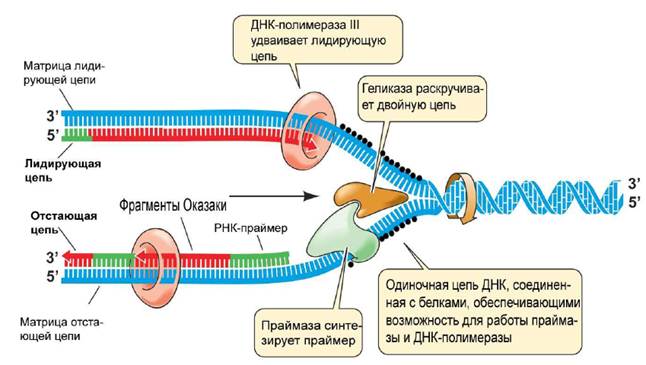
Инициация проявляется образованием репликативной вилки или локальным расщеплением цепей ДНК под воздействием фермента геликаза.
При интенсивном раскручивании цепей материнской ДНК может происходить сверхскручивание свободных концов цепей, которое устраняется ферментом топоизомераза II или гираза. Стабилизация расплетенных цепей материнской ДНК происходит при участии белка SSB (singl stand binding), который прикрепляется к одной из цепей и препятствует ее обратной рекомбинации в двойную спираль.
Синтез новой цепи ДНК происходит при помощи ферментов ДНК-полимераз. У прокариот их различают три типа (I, II, III), а у эукариот обнаружено пять типов (ά, έ, β, γ и δ).
Наиболее функционально значимой для прокариот является ДНК-полимераза III. Однако она не способна самостоятельно присоединиться к цепи ДНК и инициировать синтез новых цепей. Эту функцию выполняет фермент РНК-полимераза, при участии которого в репликационной вилке синтезируется короткий участок РНК, состоящий из 10-60 пар оснований, называемый РНК-праймером. Его структура комплементарна свободному участку ДНК.
Та цепь ДНК, где направление синтеза идет непрерывно и совпадает с направлением движения репликативной вилки, называется лидирующей, а вторая, имеющая направление синтеза, противоположное движению репликативной вилки – отстающей или запаздывающей. На лидирующей цепи матричный синтез протекает непрерывно, а на запаздывающей – в виде отдельных частей, которые называются фрагментами Оказаки.
Завершается синтез запаздывающей цепи ДНК сшиванием фрагментов Оказаки под действием фермента ДНК-лигазы. Репликация ДНК сопровождается репарацией – удалением ошибочно встроенных нуклеотидов или поврежденных участков молекулы.
Репарация ДНК (от лат. reparatio – восстановление). Может происходить под воздействием УФ-лучей, тепловых или химических факторов. При этом в цепях ДНК возникают нарушения, которые способны расстроить репликацию ДНК, что в конечном счете гибельно для клеток. Возникающие нарушения устраняются за счет многоэтапного механизма репарации. В типичном случае механизм репарации реализуется в четыре этапа.
1-й этап: специальный белковый фермент узнает и находит сайт, в котором есть нарушение;
2-й этап: эндонуклеаза выщепляет один или несколько неправильных нуклеотидов;
3-й этап: ДНК-полимераза I вставляет правильную последовательность нуклеотидов, комплементарную второй цепи ДНК;
4-й этап: ДНК-лигаза сшивает разрыв.
Рекомбинация ДНК. Для жизнедеятельности любого вида большое значение имеет не только стабильность, но и изменчивость ДНК. Нестабильная ДНК не может обеспечить приспособление организма к изменяющимся условиям существования. Рекомбинация ДНК как раз и выступает в качестве механизма адаптации организма к изменяющимся условиям среды обитания.
Различают два основных типа рекомбинации ДНК: общую и сайт-специфическую. При общей рекомбинации обмен генетическим материалом происходит с участием гомогенных (парных) нуклеотидных последовательностей ДНК. Ярким примером общей рекомбинации является процесс кроссинговера, который представляет взаимный обмен частями между ДНК, находящейся в хромосомах.
Сайт-специфическая рекомбинация происходит при интеграции ДНК фагов в значительно более длинную ДНК бактерий.
Технология и методология рекомбинантных ДНК.К началу 70-х годов 20 века были накоплены уникальные сведения о молекулах ДНК и сотнях ферментов, для которых эти молекулы являются субстратом. Если до середины 70-х годов генетический материал и его фенотипические проявления изучались в основном в пассивной манере, то в новых условиях открылись возможности для активного манипулирования этим материалом. Теперь стали выделять ДНК, вырезать из них отдельные участки, изменять и конструировать их заново, а затем вводить в геном культивируемых клеток и по фенотипическим признакам судить о генах и их функции. Рассмотрим следующие экспериментальные методы технологии рекомбинантных ДНК: расщепление ДНК, клонирование ее части, секвенирование, введение генов в вектор и затем в организм-реципиент и идентификация клеток-реципиентов.
Желая выделить ген, ДНК расщепляют посредством рестриктазы (от лат. restrictio – ограничение). Рестриктазы – это ферменты, которые вырабатываются бактериями для защиты от чужих ДНК. В настоящее время известно более 400 рестриктаз. Каждая рестриктаза узнает определенный фрагмент ДНК длиной в 4-7 нуклеотидных пар. Именно в этих местах они разрезают ДНК на части. Разрезание происходит не перпендикулярно цепям, а наискосок и одна из цепей выступает вперед на несколько нуклеотидов, образуя липкий конец. Он называется липким поскольку легко вступает во взаимодействие с другим концом, комплементарным ему.
| Рестриктазы | Участки распознавания и места разреза ДНК |
| BamI | 5' – Г –*Г – А – Т – Ц – Ц - 3' 3' – Ц – Ц – Т – А – Г - *Г - 5' |
| EcoRI | 5' – Г –*А – А – Т – Т – Ц - 3' 3' – Ц – Т – Т – А – А - *Г - 5' |
| HindIII | 5' – А –*А – Г – Ц – Т – Т - 3' 3' – Т – Т – Ц – Г – А -*А - 5' |
| HaeIII | 5' – Г –*Г – Ц – Ц - 3' 3' – Ц – Ц - Г - *Г - 5' |
| HpaII | 5' – Ц –*Ц – Г – Г - 3' 3' – Г – Г – Ц -*Ц - 5' |
| SmaI | 5' – Ц –*Ц – Ц – Г – Г – Г - 3' 3' – Г – Г – Г – Ц –Ц -*Ц - 5' |
Из фрагментов молекулы ДНК, полученных в результате обработки ее рестриктазой, можно выделить ген и при необходимости воспроизвести его путем химико-ферментативного синтеза.
В конце 70-х годов были разработаны методы секвенирования, т. е. выделения последовательности нуклеотидов. Используя химические реагенты, расщепляют ДНК на азотистые основания, и полученные образцы подвергают электрофорезу на параллельных дорожках одного геля. При этом определяется, на какой из дорожек расположена полоса. Анализируя результаты электрофореза можно установить последовательность нуклеотидов ДНК.
– Конец работы –
Эта тема принадлежит разделу:
Генетика. Курс лекций для специальности "БИОЭКОЛОГИЯ"
УО quot Витебский государственный университет... им П М Машерова quot...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Лекция 3. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ГЕНЕТИКИ
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.015 сек.
Новости и инфо для студентов