рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Образование
- /
- ПИЩЕВЫЕ ЦЕПИ И ПИЩЕВЫЕ СЕТИ
Реферат Курсовая Конспект
ПИЩЕВЫЕ ЦЕПИ И ПИЩЕВЫЕ СЕТИ
ПИЩЕВЫЕ ЦЕПИ И ПИЩЕВЫЕ СЕТИ - раздел Образование, В качестве учебного пособия Представители Разных Трофических Уровней Связаны Между Собой Односторонне Нап...
Представители разных трофических уровней связаны между собой односторонне направленной передачей биомассы в пищевые цепи. При каждом переходе на следующий трофический уровень часть доступной энергии не воспринимается, часть отдается в виде тепла, а часть расходуется на дыхание. При этом общая энергия каждый раз уменьшается в несколько раз. Следствие этого —ограниченная длина пищевых цепей. Чем короче пищевая цепь или чем ближе организм к ее началу, тем больше количество доступной энергии.
Пищевые цепи можно разделить на два основных типа: пастбищные цепи, которые начинаются с зеленого растения и идут дальше к пасущимся фитофагам, а затем к хищникам; детритные цепи, которые от мертвого органического вещества идут к детритофагам, а затем к хищникам. Пастбищные цепи, в свою очередь, объединяют пищевые цепи хищников и пищевые цепи паразитов.
Пищевые цепи хищников идут от продуцентов к травоядным, поедаемым мелкими плотоядными, а они служат пищей более крупным хищникам и т. д. По мере
продвижения по цепи хищников животные увеличиваются в размерах и уменьшаются в числе. Относительно простая и короткая пищевая цепь хищников включает консументов II порядка:
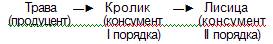
Более длинная и сложная цепь включает консументов V порядка:
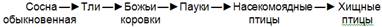
Удлинение цепи происходит благодаря участию в ней хищников.
Пищевые цепи паразитов, наоборот, ведут к организмам, которые все более уменьшаются в размерах и увеличиваются численно. Примером может служить цепь:
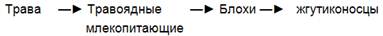
В детритных цепях консументами являются детритофаги, относящиеся к различным систематическим группам: мелкие животные, преимущественно беспозвоночные, которые живут в почве и питаются опавшей листвой, или бактерии и грибы, разлагающие органические вещества по схеме:

В большинстве случаев деятельность обеих групп детритофагов характеризуется строгой согласованностью: животные создают условия для работы микроорганизмов, разделяя трупы животных и мертвые растения на мелкие части.
Пищевые цепи, начинающиеся с зеленых растений и от мертвого органического вещества, чаще всего представлены в экосистемах совместно, но почти всегда одна из них доминирует над другой. Тем не менее, в некоторых специфических средах (например, абиссальной и подземной), где существование организмов с хлорофиллом невозможно из-за отсутствия света, сохраняются пищевые цепи только детритного типа.
Пищевые цепи не изолированы одна от другой, а тесно переплетены. Они составляют так называемые пищевые сети. Принцип образования пищевых сетей состоит в следующем. Каждый продуцент имеет не одного, а несколько консументов. В свою очередь, консументы, среди которых преобладают полифаги, пользуются не одним, а несколькими источниками питания. Для иллюстрации приведем примеры простой (рис, 9.3, а) и сложной (рис. 9.3, б) пищевых сетей.
В сложном природном сообществе те организмы, ко-
торые получают пищу от растений, занимающих первый
трофический уровень, через одинаковое число этапов, считаются принадлежащими к одному трофическому уровню. Так, травоядные занимают второй трофический уровень (уровень первичных консументов), хищники, поедающие травоядных,— третий (уровень вторичных консументов), а вторичные хищники — четвертый (уровень третичных консументов). Необходимо подчеркнуть, что трофическая классификация делит на группы не сами виды, а типы их жизнедеятельности. Популяция одного вида может занимать один или более трофических уровней, в зависимости от того, какие источники энергии эти виды используют. Точно так же любой трофический уровень представлен не одним, а несколькими видами, в результате чего цепи питания сложно переплетены.
Рассмотрим диаграмму потока энергии в простой (неразветвленной) пищевой цепи, включающей три (1-3) трофических уровня (рис. 9.4).
Для данной конкретной экосистемы энергетический бюджет оценивался следующим образом: L=3000 ккал/м2 в день, LA=1500, т.е. 50% от L, PN= 15, т.е. 1% от LA,
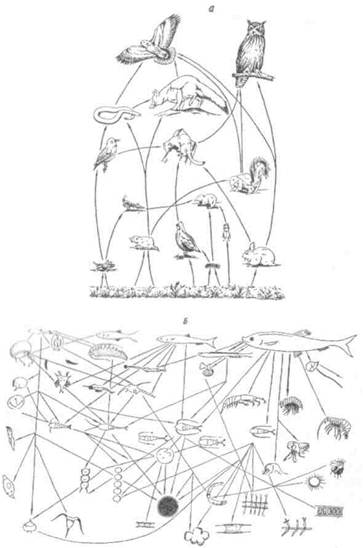
Рис. 9.3. Важнейшие связи в пищевых сетях американской прерии (а) и экосистемы северных морей для сельди (б),
а - по Риклефсу, 1979; б - из Алимова, 1989.
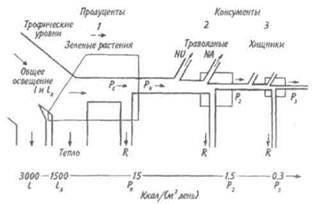
Рис. 9.4. Упрощенная диаграмма потока энергии,
показывающая три трофических уровня
в линейной пищевой цепи (по: Одум, 1975).
Последовательные потоки энергии: L — общее освещение, LA — свет,
поглощенный растительным покровом (I — поступившая или
поглощенная энергия), PG — валовая первичная продукция,
PN — чистая первичная продукция, Р— вторичная продукция (консумен-
тов), NU — не используемая энергия, NA — не ассимилированная
консументами (выделенная с экскрементами) энергия, R —энергия.
Цифры внизу — порядок потерь энергии при каждом переносе.
P2 = 1,5, т.е. 10% от PN’ , и Р3 = 0,3 ккал/м2 в день, т. е. 20% от предыдущего уровня. На первом трофическом уровне поглощается 50% падающего света, а переходит в химическую энергию пищи всего 1% поглощенной энергии. Вторичная продукция на каждом последующем трофическом уровне консументов составляет около 10% предыдущий, хотя на уровне хищников эффективность может быть выше.
Статьи поступления и расхода энергии, т.е. энергетический баланс, удобно рассмотреть с помощью универсальной модели, которая приложима к любому живому компоненту системы, будь то растение, животное, микроорганизм или же особь, популяция, трофическая группа (рис. 9.5). Не вся энергия, поступившая в биомассу (/), подвергается превращению. Часть ее (NA) не включается в метаболизм. Например, пища может пройти через пищеварительный тракт, не включаясь в мета-
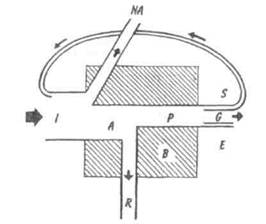
Рис. 9.5. Компоненты «универсальной» модели
потока энергии (по: Одум, 1975).
Объяснение в тексте.
болизм, а часть энергии света проходит через растения, не усваиваясь. Используемая, или ассимилируемая, часть энергии (А) тратится на дыхание (R) и продуцирование органического вещества (Р). Продукция может принимать различные формы: G – рост, или увеличение биомассы; Е – ассимилированное органическое вещество, выделяемое с экскрементами или секретируемое (простые сахара, аминокислоты, мочевина, слизь и т.д.)[4], S —запас (например, жировые накопления, которые могут быть реассимилированы позднее). Обратный путь запасенной продукции называют также «рабочей петлей», поскольку это та часть продукции, которая обеспечивает организм энергией в будущем (например, хищник использует энергию запасенных веществ для того, чтобы найти новую жертву). Оставшаяся за вычетом Е часть продукции — биомасса (В). Суммируя все статьи поступления и расхода энергии, получим: A=I-NA; P = A-R; P=G + E + S; B = P-E; B = G + S.



 Универсальную модель потока энергии можно использовать двояко. Во-первых, она может представлять популяцию какого-либо вида. В этом случае каналы притока энергии и связи данного вида с другими составляют диаграмму пищевой сети с названием отдельных видов в ее узлах (рис. 9.6). Процедура построения сетевой диаграммы включает: 1) составление схемы распределения популяций по трофическим уровням; 2) соединение их пищевыми связями; 3) определение с помощью универсальной модели ширины каналов потоков энергии; при этом наиболее широкие каналы будут проходить через популяции видов-полифагов, в данном случае через популяции поденок, мошек и комаров-дергунов (рис. 9.6).
Универсальную модель потока энергии можно использовать двояко. Во-первых, она может представлять популяцию какого-либо вида. В этом случае каналы притока энергии и связи данного вида с другими составляют диаграмму пищевой сети с названием отдельных видов в ее узлах (рис. 9.6). Процедура построения сетевой диаграммы включает: 1) составление схемы распределения популяций по трофическим уровням; 2) соединение их пищевыми связями; 3) определение с помощью универсальной модели ширины каналов потоков энергии; при этом наиболее широкие каналы будут проходить через популяции видов-полифагов, в данном случае через популяции поденок, мошек и комаров-дергунов (рис. 9.6).
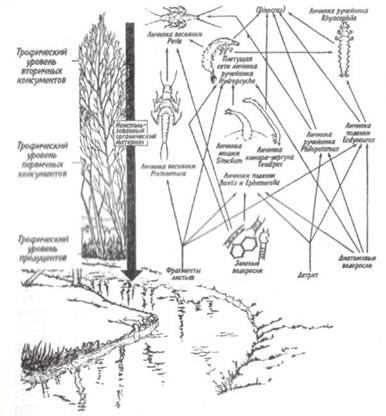 Рис. 9.6. Фрагмент пищевой сети пресноводного водоема.
Рис. 9.6. Фрагмент пищевой сети пресноводного водоема.
Во-вторых, универсальная модель потока энергии может представлять определенный энергетический уровень. В этом варианте прямоугольники биомассы и каналы потоков энергии представляют все популяции, поддерживаемые одним источником энергии. Обычно пищей лисам служат частично растения (плоды и т. д.), частично травоядные животные (зайцы, полевые мыши и др.). Если мы хотим подчеркнуть аспект внутрипопуляционной энергетики, то всю популяцию лис необходимо изобразить одним прямоугольником. Если же требуется распределить метаболизм популяции лис на два трофических уровня соответственно пропорции растительной и животной пищи, то следует построить два или несколько прямоугольников.
Зная универсальную модель потока энергии, можно определить отношения величин энергетического потока в разных точках пищевой цепи. Выраженные в процентах, эти отношения называют экологической эффективностью. В зависимости от задач исследования эколог изучает те или иные группы экологических эф-фективностей. Важнейшие из них рассматриваются ниже.
Первая группа энергетических отношений: В/R и P/R. Часть энергии, идущая на дыхание, т.е. на поддержание структуры биомассы, велика в популяциях крупных организмов (люди, деревья и др.) При сильном стрессе R возрастает. Величина Р значительна в активных популяциях мелких организмов, например бактерий и водорослей, а также в системах, получающих энергию извне.
Вторая группа отношений: А/I и Р/А. Первое из них называется эффективностью ассимиляции, второе —эффективностью роста тканей. Эффективность ассимиляции варьирует от 10 до 50% и больше. Она может быть либо очень мала, как в случае использования энергии света растениями или при ассимиляции пищи животными-детритофагами, либо очень велика, как в случае ассимиляции пищи животными или бактериями, питающимися высококалорийной пищей, например сахарами или аминокислотами.
Эффективность ассимиляции у растительноядных животных соответствует питательным свойствам их пищи: она достигает 80% при поедании семян, 60% —молодой листвы, 30-40% —более старых листьев и 10-20% и даже менее при поедании древесины, в зависимости от степени ее разложения. Пища животного происхождения переваривается легче, чем растительная. Эффективность ассимиляции у хищных видов составляет 60-90% потребленной пищи, причем виды, поедающие насекомых, стоят на нижней ступени этого ряда, а питающиеся мясом и рыбой — на верхней. Причина такого положения заключается в том, что жесткий хитиновый наружный скелет, на который приходится значительная часть массы тела у многих видов насекомых, не переваривается. Это снижает эффективность ассимиляции у животных, которые питаются насекомыми.
Эффективность роста тканей также широко варьирует. Наибольших значений она достигает в тех случаях, когда организмы мелкие и условия среды, в которой они обитают, не требуют больших затрат на поддержание оптимальной для роста организмов температуры.
И, наконец, третья группа энергетических отношений: Р/В.
В тех случаях, когда Р оценивается как скорость, Р/В представляет собой отношение продукции в конкретный момент времени к биомассе: Р/В = В/(ВТ) = Т-1, где Т - время. Если рассчитывается интегральная продукция за некоторый промежуток времени, значение отношения Р/В определяется с учетом средней за этот же отрезок времени биомассы. В этом случае отношение Р/В - величина безразмерная; она показывает, во сколько раз продукция больше или меньше биомассы. Отношение продуктивности к биомассе можно рассматривать как внутри одного трофического уровня, так и между соседними.
Сравнивая продуктивность Pt и биомассу Bt внутри одного трофического уровня (t), отметим S-образный характер изменения Pt в определенном диапазоне изменений Bt. Например, на первом трофическом уровне продукция увеличивается сначала медленно, так как невелика поверхность листьев, затем быстрее и при большой плотности биомассы — опять медленно, поскольку
фотосинтез в условиях значительного затенения листьев нижних ярусов ослабляется. На втором и третьем трофических уровнях при очень малом и при очень большом числе животных на единицу площади отношение продуктивности к биомассе снижается, главным образом из-за уменьшения рождаемости.
Отношение продуктивности предыдущего трофического уровня (Рt-1) к биомассе настоящего (Bt) определяется тем, что фитофаги, выедая часть растений, тем самым способствуют ускорению их прироста, т. е. фитофаги своей деятельностью содействуют продуктивности растений. Аналогичное влияние оказывают на продуктивность консументов I порядка хищники, которые, уничтожая больных и старых животных, способствуют повышению коэффициента рождаемости фитофагов.
Наиболее проста зависимость продуктивности последующего трофического уровня (Pt+1) от биомассы настоящего (Вt). Продуктивность каждого последующего трофического уровня возрастает при росте биомассы предыдущего.Отношение Рt+1/Bt показывает, в частности, от чего зависитвеличина вторичной продукции, а именно отвеличины первичной продукции, длины пищевой цепи, природы и величины энергии, привносимой извне в экосистему.
Приведенные рассуждения позволяют заметить, что на энергетические характеристики экосистемы определенное влияние оказывают размеры особей. Чем мельче организм, тем выше его удельный метаболизм (на единицу массы) и, следовательно, меньше биомасса, которая может сохраняться на данном трофическом уровне. И наоборот, чем крупнее организм, тем больше биомасса на корню. Так, «урожай» бактерий в данный момент будет гораздо ниже «урожая» рыбы или млекопитающих, хотя эти группы использовали одинаковое количество энергии. Иначе обстоит дело с продуктивностью. Поскольку продуктивность — это скорость прироста биомассы, то преимуществами здесь обладают мелкие организмы, которые благодаря более высокому уровню
метаболизма имеют более высокие темпы размножения и обновления биомассы, т. е. более высокую продуктивность.
– Конец работы –
Эта тема принадлежит разделу:
В качестве учебного пособия
На сайте allrefs.net читайте: В качестве учебного пособия.
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: ПИЩЕВЫЕ ЦЕПИ И ПИЩЕВЫЕ СЕТИ
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.025 сек.
Новости и инфо для студентов