ТРОФИЧЕСКАЯ СТРУКТУРА ЭКОСИСТЕМЫ - раздел Образование, В качестве учебного пособия В Результате Рассеяния Энергии В Пищевых Цепях И Благодаря Такому Фактору, Ка...
В результате рассеяния энергии в пищевых цепях и благодаря такому фактору, как зависимость метаболизма от размера особей, каждое сообщество приобретает определенную трофическую структуру, которую можно выразить либо числом особей на каждом трофическом уровне, либо урожаем на корню (на единицу площади), либо количеством энергии, фиксируемой на единице площади за единицу времени на каждом последующем трофическом уровне. Графически это можно представить в виде пирамиды, основанием которой служит первый трофический уровень, а последующие образуют этажи и вершину пирамиды. Различают три основных типа экологических пирамид — пирамиды чисел, биомассы и энергии.
Пирамида чисел, или распределение особей по трофическим уровням, зависит от следующих факторов:
1. В любой экосистеме мелкие животные численно превосходят крупных и размножаются быстрее.
2. Для всякого хищного животного существуют нижний и верхний пределы размеров их жертв. Верхний предел определяется тем, что хищник не в состоянии одолеть животное, намного превышающее по размеру его собственное тело, нижний предел – тем, что при слишком малом размере добычи, охота на нее теряет для хищников какой-либо смысл. Мелкую добычу хищникам пришлось бы ловить в таких огромных количествах, что это оказалось бы почти невозможным либо ввиду ее недостатка, либо нехватки времени. Поэтому для хищников каждого вида, как правило, пищей служат жертвы оптимального размера.
Для поддержания жизни одного льва требуется 50 зебр в год. На Фолклендских островах кулик-сорока на скалах во вре-
мя отлива добывает брюхоногого моллюска «блюдечко», но может справиться только с особями среднего размера; моллюски, размер которых превышает 45 мм, от него ускользают.
Однако из этого правила есть исключения. Волки, охотясь сообща, могут убивать жертву более крупную, чем они сами, например оленей. Пауки и змеи, обладая свойством выделять яд, убивают крупных животных. Единственным же видом, способным использовать в пищу животных любой величины, является человек.
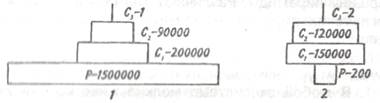
В случае пастбищных пищевых цепей леса, когда продуцентами служат деревья, а первичными консументами — насекомые, уровень первичных консументов численно богаче особями уровня продуцентов. Таким образом, пирамиды чисел могут быть обращенными. Для примера на рис. 9.7 приведены пирамиды чисел экосистем степи и леса умеренной зоны.
Рис. 9.7. Пирамида чисел для степи летом (1)
и для леса умеренной зоны летом (2) (из Одума, 1975).
Р — продуценты, С — консументы.
Пирамиды биомассы представляют более фундаментальный интерес, так как в них устранен «физический» фактор и четко показаны количественные соотношения биомасс. Если организмы не слишком сильно различаются по размеру, то, обозначив на трофических уровнях общую массу особей, можно получить ступенчатую пирамиду. Но если организмы низших уровней в среднем мельче организмов высших уровней, то имеет место обращенная пирамида биомассы. Например, в экосистемах с очень мелкими продуцентами и крупными консументами общая масса последних может быть в любой
данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений.
1. Наземные и мелководные экосистемы, где продуценты крупные и живут сравнительно долго, характеризуются относительно устойчивыми пирамидами с широким основанием и узкой вершиной. На форму пирамиды подобных экосистем влияет также возраст сообщества. В недавно возникших сообществах отношение биомассы консументов к биомассе продуцентов обычно меньше, чем в зрелых (т. е. вершина пирамиды будет более узкой). Объясняется это тем, что консументы наземных и мелководных сообществ имеют более сложные жизненные циклы и более «изощренные» требования к местообитанию (например, им нужны особые укрытия), чем зеленые растения. Поэтому животным популяциям может требоваться больше времени для максимального развития.
2. В открытых и глубоких водах, где продуценты невелики по размеру и имеют короткий жизненный цикл, пирамида биомассы может быть обращенной. Общий урожай на корню здесь, как правило, меньше, чем в наземных или мелководных сообществах, даже если количество фиксируемой за год энергии в обоих случаях одинаково.
3. В озерах и прудах, где равное значение как продуценты имеют и крупные прикрепленные растения, и микроскопические водоросли, пирамида урожая на корню будет иметь промежуточный вид.
Пирамида энергии из трех рассматриваемых типов экологических пирамид дает наиболее полное представление о функциональной организации сообщества, так как число и масса организмов, которые могут существовать на каждом трофическом уровне в тех или иных условиях, зависят не от количества фиксированной энергии, имеющейся в данное время на предыдущем уровне, а от скорости продуцирования пищи. В противоположность пирамидам чисел и биомассы, отражающим статику системы, т. е. характеризующим количество организмов или их биомассу в данный момент, пирамида энергии отра-
жает скорость прохождения массы пищи через пищевую цепь. На форму этой пирамиды не влияют изменения размеров особей и интенсивности их метаболизма, и если учтены все источники энергии, то пирамида всегда будет иметь «канонический» вид, как это диктуется вторым законом термодинамики.
Для того чтобы оценить достоинства модели пирамиды энергии, сравним пирамиды биомассы и энергии одной из немногих экосистем, для которых известны все компоненты сообщества, включая редуцентов, — экосистемы ручьев Силвер-Спрингс во Флориде (рис.9.8).
Продуценты в данной экосистеме представлены стрелолистом и прикрепленными водорослями. Среди кон-
Рис. 9.8. Пирамиды биомассы (1) и энергии (2) в системе
ручьев Силвер-Спрингс во Флориде (из Одума, 1975).
S —редуценты. Р, С — см. рис. 9.7.
сументов I порядка большое количество насекомых, брюхоногих моллюсков, растительноядных рыб и черепах. Различные виды рыб и хищные насекомые занимают следующий трофический уровень, а окунь, паразиты и панцирная щука - самый верхний уровень. Редуценты не только разлагают растительный материал, но уничтожают и органическое вещество других уровней: на пирамиде биомассы им соответствует линия, опирающаяся на первый трофический уровень. Биомасса бактерий и грибов очень мала по сравнению с их ролью в энергетическом потоке сообщества. Поэтому в пирамиде чисел значение редуцентов сильно преувеличено, а в пирамиде биомассы сильно преуменьшено. Исходя из данного положения, сформулируем «экологическое правило»: данные по численности приводят к преувеличению зна-
чения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов.
Следовательно, эти критерии непригодны для сравнения функциональной роли популяций, сильно различающихся по значению отношения интенсивности метаболизма к размеру особей, хотя, как правило, биомасса более надежный критерий, чем численность. В то же время поток энергии служит более подходящим показанием при сравнении компонента с любым другим и экосистемы в целом с соседней экосистемой.
Поток энергии в экосистеме представляет интерес не только для эколога, но и для каждого из нас. Объясняется это тем, что, уяснив основные законы превращения энергии, мы лучше поймем, почему природные экосистемы функционируют именно так, как они функционируют. Не менее важно и то, что такое знание поможет нам определить границы, за которыми наше воздействие на окружающую среду принесет ей непоправимый ущерб.
На сайте allrefs.net читайте: В качестве учебного пособия.
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
ТРОФИЧЕСКАЯ СТРУКТУРА ЭКОСИСТЕМЫ
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
К первому и второму изданиям
В современном мире экологические проблемы по своей общественной значимости вышли на одно из первых мест, оттеснив даже опасность ядерной войны. Бурное развитие хозяйственной деятельности людей при
К третьему изданию
Со времени опубликования второго издания учебного пособия «Краткий курс общей экологии» прошло всего 2 года. Срок по всем меркам очень малый. И, тем не менее, за это время в нашей жизни зародились
ЭКОЛОГИЯ - ЗАДАЧИ И ПЕРСПЕКТИВЫ
Подобно всем другим областям знания, экология развивалась непрерывно, но неравномерно. Труды Гиппократа, Аристотеля и иных древнегреческих философов содержат сведения явно экологического характер
ЭКОЛОГИЯ КАК НАУКА
Как самостоятельная наука экология сформировалась приблизительно к 1900 г. Термин «экология» был предложен немецким биологом Эрнстом Геккелем в 1869 г. Следовательно, это сравнительно молодая наук
ПРОБЛЕМЫ, ИЗУЧАЕМЫЕ ЭКОЛОГИЕЙ
Имеется несколько кардинальных проблем, в которых фокусируются основные направления и разделы современной экологии. Успехи в их решении в значительной мере определяют прогресс всей экологии, и поэ
ПРИНЦИП ДЕЙСТВИЯ ЭКОЛОГИЧЕСКОГО ФАКТОРА
Понятие условий экология заменила понятием фактора. Любой организм в среде своего обитания подвергается воздействию самых разнообразных климатических, эдафических и биотических факторов. «Экологи
РЕАКЦИЯ ОРГАНИЗМОВ НА ОДНОВРЕМЕННОЕ ДЕЙСТВИЕ НЕСКОЛЬКИХ ФАКТОРОВ
В природных условиях организм подвержен воздействию многочисленных факторов. Если действие одного фактора описывается соответствующей функцией, то одновременное действие двух факторов можно изобр
СРЕДА И ДЕЙСТВИЕ ФАКТОРОВ СРЕДЫ
Часть природы, окружающая организмы и оказывающая на них прямое или косвенное воздействие, часто обозначают как «среда». Из среды особи получают все необходимое для жизни и в нее же выделяют проду
ВИТАЛЬНОЕ И СИГНАЛЬНОЕ ДЕЙСТВИЕ ФАКТОРОВ
Если положить в основу классификации экологических факторов эффект, который вызван их воздействием, а именно этот принцип был использован в преды-
дущей классификации, то более рацион
УРОВЕНЬ ОСОБЕЙ
На уровне особи абиотические факторы оказывают влияние прежде всего на поведение животных, изменяют трофические отношения со средой и характер метаболизма, воздействуют на способность к размножен
ПОПУЛЯЦИОННЫЙ УРОВЕНЬ
На уровне популяции абиотические факторы воздействуют на такие параметры, как рождаемость, смертность, средняя продолжительность жизни особи, скорость роста популяции и ее размеры, нередко являя
ВИДОВОЙ УРОВЕНЬ
На уровне вида абиотические условия среды обитания нередко выступают как факторы, определяющие и ограничивающие географическое распространение. Кроме того, разнообразие климата, физических и хими
УРОВЕНЬ ЭКОЛОГИЧЕСКИХ СИСТЕМ
Наше знакомство с общими закономерностями воздействия абиотических факторов было бы неполным без рассмотрения специфики проявления его на уровне экосистем. Особенности влияния экологических факто
ЭКОЛОГИЧЕСКАЯ РОЛЬ ФАКТОРОВ ПИТАНИЯ
Многие экологические термины можно объединить в пары, члены которых имеют противоположный смысл. Продуцент и консумент относятся к различным сторонам активности одного и того же орга
ПИЩА КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР
Основным способом питания растения является фотосинтез, в процессе которого под действием солнечной энергии происходит восстановление углекислого газа до углеводов (СН20):
ПИЩЕВЫЕ РЕЖИМЫ И ПИЩЕВАЯ СПЕЦИАЛИЗАЦИЯ ЖИВОТНЫХ
Для обсуждения роли количества и качества питательных веществ в жизнедеятельности животных рассмотрим их пищевые режимы и пищевые специализации. Под пищевым режимом понимается своеобразие, природа
ГОМОТИПИЧЕСКИЕ РЕАКЦИИ
Под гомотипическими реакциями понимают взаимодействия между особями одного вида. Жизнедеятельность животных и растений при совместном обитании в значительной степени зависит от численности и плот
ПРИНЦИП КОНКУРЕНТНОГО ИСКЛЮЧЕНИЯ. ЭКОЛОГИЧЕСКАЯ ДИВЕРСИФИКАЦИЯ
Решающее значение в природе имеет межвидовая конкуренция, поскольку она в большей степени, чем другие гетеротипические реакции, определяет роль видов в экосистемах. Межвидовая конкуренция — это та
ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Объектом изучения демоэкологии, или популяционной экологии, служит популяция. Ее определяют как группу организмов одного вида (внутри которой особи могут обмениваться генетической информацией), зан
ЭКСПОНЕНЦИАЛЬНАЯ И ЛОГИСТИЧЕСКАЯ КРИВЫЕ РОСТА
Численность популяции определяется в основном двумя противоположными явлениями — рождаемостью и смертностью. Пусть N — численность популяции. Тогда отношение dN/dt означает мгновенну
СВОЙСТВА ПОПУЛЯЦИЙ
От чего же зависят значения коэффициентов рождаемости и смертности? От очень многих факторов, действующих на популяцию извне, а также от собственных ее свойств. Объективный показатель способности
КОЛЕБАНИЯ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Когда популяция прекращает расти, ее плотность обнаруживает тенденцию к флуктуациям относительно верхнего асимптотического уровня роста. Такие флуктуации могут возникать либо в результате изменен
ФАКТОРЫ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Известно три типа зависимости численности популяции от ее плотности (рис. 7.10). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко ра
КОНЦЕПЦИЯ ЭКОСИСТЕМЫ
Первые организмы на Земле были гетеротрофами. Они быстро исчерпали бы себя, если бы не появились автотрофы. При наличии этих групп организмов уже возможен примитивный круговорот веществ:
ГОМЕОСТАЗ ЭКОСИСТЕМЫ
Сложившаяся исторически экосистема не должна рассматриваться просто как сумма слагаемых, т. е. сочетание отдельных входящих в ее состав организмов. Это система, сохраняющая устойчивость при относ
ПОТОК ЭНЕРГИИ В ЭКОСИСТЕМЕ
Важнейшая термодинамическая характеристика экосистемы—ее способность создавать и поддерживать высокую степень внутренней упорядоченности, т. е. состояние с низкой энтропией[2]. Система обладает н
ПИЩЕВЫЕ ЦЕПИ И ПИЩЕВЫЕ СЕТИ
Представители разных трофических уровней связаны между собой односторонне направленной передачей биомассы в пищевые цепи. При каждом переходе на следующий трофический уровень часть доступной энерг
ОБМЕННЫЙ И РЕЗЕРВНЫЙ ФОНДЫ
Известно, что из более 90 химических элементов, встречающихся в природе, 30-40 необходимы живым организмам. Некоторые элементы, такие как углерод, водород и азот, требуются в больших количествах, д
БЛОЧНАЯ МОДЕЛЬ КРУГОВОРОТА
Существуют различные способы изображения биогеохимических круговоротов. Выбор способа зависит от особенностей биогеохимического цикла того или иного элемента. При обсуждении круговорота кислорода
ВИДОВАЯ СТРУКТУРА БИОТИЧЕСКОГО СООБЩЕСТВА
Структура биотического сообщества слагается из нескольких показателей.
Обилие — число особей на единицу площади или объема.
Частота — отношение числа особей одного
ВНУТРЕННЯЯ ОРГАНИЗАЦИЯ БИОТИЧЕСКОГО СООБЩЕСТВА
Облик биотического сообщества определяется не только разнообразием видов и другими показателями, которые отражают связи между видами, входящими в состав биотического сообщества. Функционирование со
РАЗВИТИЕ И ЭВОЛЮЦИЯ ЭКОСИСТЕМЫ
Одно из основных свойств экосистем - их динамизм. Наблюдение над полем, заброшенным на несколько лет, показывает, что его последовательно завоевывают сначала многолетние травы, затем кустарники и,
ЭКОЛОГИЧЕСКАЯ СУКЦЕССИЯ
Автогенные изменения называют развитием экосистемы, или экологической сукцессией. При определении экологической сукцессии следует учитывать три момента:
1. Сукцессия происходит под дейст
ПЕРВИЧНАЯ И ВТОРИЧНАЯ СУКЦЕССИИ. ПОНЯТИЕ КЛИМАКСА
Экологическая сукцессия протекает через ряд этапов, при этом биотические сообщества сменяют друг друга. Замещение видов в сукцессии вызвано тем, что популяции, стремясь модифицировать окружающую с
ЭВОЛЮЦИЯ ЭКОСИСТЕМЫ
Вопрос о том, как эволюционируют экосистемы, очень важен, поскольку его решение — ключ к пониманию существующего разнообразия сообществ живых организмов на нашей планете, смены флор и фаун в ходе
ХОРОЛОГИЧЕСКИЙ АСПЕКТ ИЗУЧЕНИЯ ЭКОСИСТЕМЫ
Выделение (разграничение) экосистем в природе часто оказывается далеко не простой задачей. Прежде всего, не существует единой точки зрения по поводу минимальной размерной единицы экосистемы. Нек
ПРИНЦИПЫ РАЗГРАНИЧЕНИЯ БИОГЕОЦЕНОЗОВ
При разграничении экосистем возникает проблема выбора одного из характерных признаков, которые подразделяют на физиономические, таксономические и экологические. На основе физиономических признако
ИЕРАРХИЧЕСКИЙ РЯД ЭКОСИСТЕМ
Установив минимальную размерную единицу экосистемы — биогеоценоз, можно построить иерархический ряд экосистем (рис. 13.1). Применяя термин «биогеоценоз» в этом смысле, мы сохраняем знак равенства
БИОСФЕРА
Понятие биосферы вошло в науку случайно. Более 100 лет назад, в 1875 г., австрийский геолог Эдуард Зюсс, говоря о различных оболочках земного шара, впервые упот-
ребил этот термин в по
Дополнительная
Алимов А. Ф. Введение в продукционную гидробиологию. Л., 1989.
Гиляров А. М. Популяционная экология. М., 1990. 191 с.
Лархер В. Экология растений. М., 1978. 185 с.
Леме Ж
Хотите получать на электронную почту самые свежие новости?
Подпишитесь на Нашу рассылку
Наша политика приватности обеспечивает 100% безопасность и анонимность Ваших E-Mail


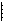 данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений.
данный момент выше общей массы продуцентов. Для пирамид биомассы можно сделать несколько обобщений.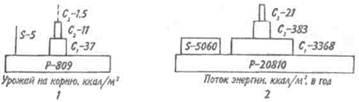


 чения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов.
чения мелких организмов, а данные по биомассе — к преувеличению роли крупных организмов.





Новости и инфо для студентов