рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Образование
- /
- Упаковка ДНК в хромосомах
Реферат Курсовая Конспект
Упаковка ДНК в хромосомах
Упаковка ДНК в хромосомах - раздел Образование, Тема ХРОМОСОМНАЯ ТЕОРИЯ Молекулы Днк В Эукариотических Клетках Очень Велики. Так, Длина Молекул Днк,...
Молекулы ДНК в эукариотических клетках очень велики. Так, длина молекул ДНК, выделенных из клеток человека, достигает нескольких сантиметров. Принято считать, что каждая эукариотическая хромосома содержит одну — единственную непрерывную молекулу ДНК. Учитывая видовое количество хромосом у млекопитающих, можно сказать, что в среднем у них на интерфазное ядро приходится около 2 м ДНК, находящейся в сферическом ядре диаметром менее 10 мкм. При этом в ядре должен сохраняться определенный порядок расположения молекул ДНК, чтобы обеспечить ее упорядоченное функционирование.
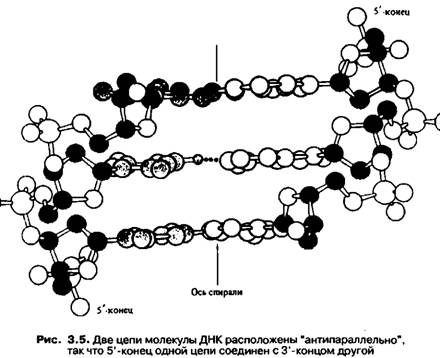


Молекулы ДНК в ядрах эукариотических клеток всегда находятся в комплексе с белками в составе хроматина, который образуется из хромосом после окончания деления ядер в результате сложного процесса раскручивания (деспирализации) хромосом.
На долю белков приходится около 60% сухого веса хроматина. Белки в его составе очень разнообразны. Обычно их разделяют на две группы: гистоны и негистоновые белки. Именно гистоны, характерные только для эукариотических клеток, осуществляют первые этапы упаковки ДНК, очень схожие у большинства изученных объектов
На долю гистонов приходится до 80% всех белков хроматина. Их взаимодействие с ДНК происходит за счет ионных связей и не зависит от последовательности нуклеотидов в составе молекулы ДНК. Гистоны не отличаются большим разнообразием. Это глобулярные белки, представленные 5-7 типами молекул. Наиболее известны следующие классы гистонов: HI, Н2А, Н2В, НЗ и Н4. Их основные свойства определяются относительно высоким содержанием основных аминокислот: лизина и аргинина (рис. 3.7). Положительные заряды на аминогруппах указанных аминокислот обеспечивают электростатическую связь гистонов с отрицательными зарядами на фосфатных группах ДНК. Из всех ядерных белков гистоны изучены наиболее хорошо. Их молекулярная масса относительно невелика (максимальная — у гистона НЗ — 153 тыс. дальтон). Практически у всех эукариот они обладают сходными свойствами и подразделяются на одни и те же классы. Из исследованных эти белки наиболее консервативны: их аминокислотные последовательности близки даже у отдаленных видов. Исключение составляют гистоны HI, для которых характерны значительные межвидовые и межтканевые вариации
В процессе жизнедеятельности клеток гистоны могут подвергаться посттрансляционным модификациям, что изменяет их свойства и способность связываться с ДНК. Гистоны синтезируются в цитоплазме, переносятся в ядро и связываются с ДНК во время ее репликации в S-периоде клеточного цикла. Включившиеся в хроматин гистоны очень стабильны и имеют низкую скорость обмена.
Присутствие гистонов во всех эукариотических клетках, их сходство даже у очень отдаленных видов, обязательность в составе хромосом и хроматина — все это говорит о чрезвычайно важной роли этих белков в жизнедеятельности клеток. Этапным событием в изучении упаковки ДНК в составе хроматина стало открытие нуклеосом частиц, в которых происходит первый этап упаковки ДНК в хроматине. Сердцевина нуклеосомы всегда консервативна, содержит восемь молекул: по две молекулы гистонов Н4, НЗ, Н2А, Н2В. По поверхности сердцевины располагается участок ДНК из 146 нуклеотидных пар, образующий 1,75 оборота вокруг сердцевины. Небольшой участок ДНК остается несвязанным с сердцевиной, он называется линкером (рис. 3.8). В разных объектах линкерный участок может варьировать от 8 до 114 нуклеотидных пар на нуклеосому
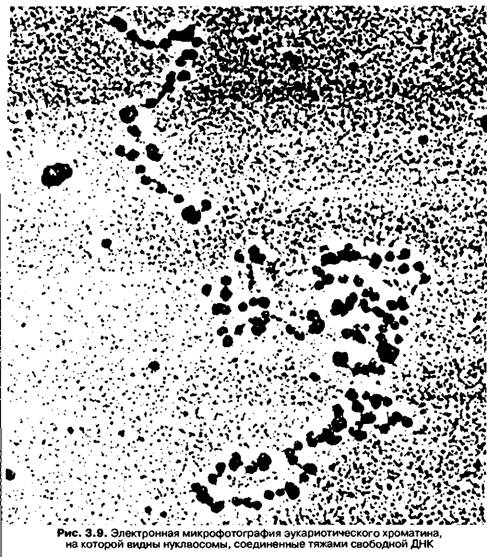
Рассчитано, что на весь гаплоидный геном человека (3 х 109 пар оснований) приходится 1,5 х 107 нуклеосом. Общий вид хроматина, представленного молекулой ДНК, упакованной с помощью нуклеосомных структур, можно сравнить с бусами на нитке (рис. 3.9). Нуклеосомы способны к самосборке при наличии в пробирке ДНК и гистонов в определенном соотношении. Первый нуклеосомный уровень компактизации ДНК увеличивает плотность упаковки ДНК в 6-7раз.

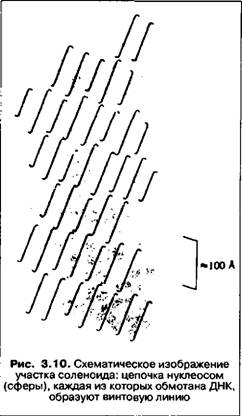
В следующий этап упаковки нуклеосомная структура хроматина вовлекается с помощью гистона HI, который связывается с линкернои частью ДНК и поверхностью нуклеосомы. Благодаря сложному взаимодействию всех компонентов возникает упорядоченная структура спирального типа, которую часто называют соленоидом (рис. 3.10). Она повышает компактность ДНК еще в 40 раз. Поскольку соленоидная структура имеет сниженную способность связываться с белками, обеспечивающими транскрипцию, то считается, что этот уровень компактизации ДНК может играть роль фактора, инактивирующего гены. Некоторые авторы рассматривают соленоидную структуру как один из возможных вариантов упаковки хроматина
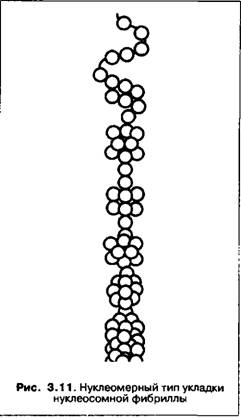
с помощью гистона HI и полагают вероятным существование и других морфологических вариантов, например, нуклеомер, или сверхбусин (рис. 3.11).
Более высокие уровни компактизации ДНК в хроматине связаны с негистоновыми белками. На их долю приходится около 20% всех белков хроматина. Эту сборную группу белков отличает широкий спектр свойств и функций. Всего фракция негистоновых белков объединяет около 450 индивидуальных белков, свойства и конкретные функции которых еще не достаточно изучены. Выяснено, что некоторые из них специфично связываются с определенными участками ДНК, в результате чего фибриллы хроматина в местах связывания ДНК с не гистоновыми белками образуют петли. Таким образом, более высокие уровни упаковки ДНК в составе хроматина обеспечиваются не спирализацией нитей хроматина, а образованием поперечной петлистой структуры вдоль хромосомы (рис. 3.12). На всех указанных этапах компактизации ДНК хроматин представлен в активной форме, в нем происходит транскрипция, синтез всех типов молекул РНК. Такой хроматин называют эухроматином. Дальнейшая упаковка хроматина ведет к переходу его в неактивное состояние с образованием гетерохроматина
Этот процесс связан со спирализацией групп петель и образованием из фибрилл хроматина розеткоподобных структур, которые обладают оптической и электронной плотностью и называются хромомерами (рис. 3.12). Предполагается, что вдоль хромосомы расположено большое количество хромомер, соединенных между собой в единую структуру участками хроматина с пуклеосомной или соленоидной упаковкой ДНК. Каждая пара гомологичных хромосом имеет свой хромомерный рисунок, который можно выявить с помощью специальных методов окрашивания при условии сиирализации хроматина и перехода его в состояние хромосом.
Петельно-розеточная структура хроматина обеспечивает не только упаковку ДНК, но и организует функциональные хромосом, поскольку в своих основаниях петли ДНК связаны с негистоновыми белками, в состав которых могут входить ферменты репликации, обеспечивающие удвоение ДНК, и ферменты транскрипции, благодаря которым происходит синтез всех типов РНК.
Участки ДНК, упакованные в виде гетерохроматина, могут иметь двоякую природу. Различают два типа гетерохроматина: факультативный и конститутивный (структурный). Факультативный гетерохроматин представляет собой участки генома, временно инактивированные в тех или иных клетках. Примером такого хроматина служит половой гетерохроматин инактивированной Х-хромосомы в соматических клетках женщин. Структурный гетерохроматин во всех клетках постоянно находится в неактивном состоянии и, вероятно, выполняет структурные или регуляторные функции.

– Конец работы –
Эта тема принадлежит разделу:
Тема ХРОМОСОМНАЯ ТЕОРИЯ
Хххххххх хххххххх хххххх Тема ХРОМОСОМНАЯ ТЕОРИЯ... Занятие Генные мутации... Занятие Хромосомные и геномные мутации...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Упаковка ДНК в хромосомах
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.015 сек.
Новости и инфо для студентов