рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Образование
- /
- Организация генетического материала в хромосомах человека
Реферат Курсовая Конспект
Организация генетического материала в хромосомах человека
Организация генетического материала в хромосомах человека - раздел Образование, Тема ХРОМОСОМНАЯ ТЕОРИЯ Общая Организация Хромосом Человека Традиционна: В Метафазе Хромосома Состо...
Общая организация хромосом человека традиционна: в метафазе хромосома состоит из двух сестринских хроматид, соединенных между собой в районе первичной перетяжки (центромеры). Центромера делит хроматиду на два плеча. Плечи могут быть равными, тогда хромосома называется метацентрической. Если одно плечо немного короче другого, то хромосомы именуются субметацентрическими. В нескольких нарах хромосом человека одно плечо сильно короче другого, такие хромосомы носят название акроцентрических (рис. 3.13). Тонкая морфология хромосом зависит от фазы митоза. Наиболее сильно спирализованы хромосомы в мета- и анафазе.
Вместе с морфологией хромосом изменяется в ходе митозе и морфология центромеры. Наиболее четко центромера выражена в виде более тонкого и светлого участка хромосомы к концу профазы. Центромеры выполняют в хромосомах очень важные функции. Они соединяют две сестринские хроматиды, велика также их роль в организации веретена деления. В районе центромеры в профазе формируется особая белковая структура, имеющая сродство к белкам микротрубочек веретена деления
Микротрубочки веретена деления соединяются с хроматидами в районе центромеры так, что на один центромерный район может приходиться более десяти микротрубочек. Очень важным моментом в прохождении митоза является синхронность одновременного разделения всех хромосом на две хроматиды. Считается, что ведущая роль в регуляции этого процесса также принадлежит центромерам.
Рассматривая общую морфологию хромосом, нельзя обойти вниманием их концевые участки, называемые теломерами. Концы хромосом — теломеры, имеют особенности в первичной и третичной структурах, но об этом речь пойдет несколько позже. Сначала ознакомимся с функциями теломерных районов. Когда деление клетки закончено и формируются новые клеточные ядра, то с помощью теломер хромосомы прикрепляются к внутренней ядерной мембране, в результате чего каждая хромосома в деспирализованном состоянии занимает в ядре строго определенное место. Помимо этого теломерные районы предотвращают слипание хромосом своими концами и препятствуют образованию дицентриков хромосом с двумя центромерами, наличие которых свидетельствует о патологических картинах митоза
Одновременно теломеры стабилизируют хромосомы, защищая их от деградации клеточными нуклеазами — ферментами, катализирующими гидролиз всех незащищенных ДНК или их фрагментов. В последнее время стало известно еще одно назначение теломерных концов: благодаря им происходит полное завершение редупликации хромосом при подготовке клетки к делению. Среди ферментов, участвующих в удвоении ДНК, помимо уже известных ДНК-нолимеразы, ДНК-лигазы, геликазы, топоизомеразы, а также стабилизирующих белков, особое внимание следует уделить теломеразе, которая помогает завершить репликацию ДНК на отстающей цепи.

Принято считать, что каждая хроматида содержит одну из двух идентичных дочерних молекул ДНК, образующихся в процессе репликации. Молекула ДНК представляет собой непрерывную сверхскрученную двойную спираль, простирающуюся по всей длине хроматиды. Функционально эта нить подразделяется на большое число отрезков, соответствующих отдельным генам. Каждый ген несет информацию о первичной структуре отдельной полипиптидной цепи, рибосомной РНК, транспортной РНК или выполняет регуляторную функцию. Кроме того, в составе непрерывной нити ДНК, наряду со смысловыми генами, находятся многократно повторяющиеся одинаковые или сходные по составу нуклеотидные последовательности, выполняющие, вероятно, регуляторные или структурные функции.
Информация о первичной структуре полипептидов (последовательности аминокислот в них) записана в ДНК в виде трехбуквенного кода, составленного из первых букв названий четырех азотистых оснований, входящих в состав ДНК (АТГЦ). Каждой аминокислоте соответствует определенный триплет из трех соседних нуклеотидов. Например, аминокислоте фенилаланин в ДНК соответствует кодон AAA, а аминокислоте серии — АГА.
В пределах одного гена, который кодирует полипептид, участок молекулы ДНК подразделяется на функционально различные единицы (рис. 3.14). Отличительная черта строения многих генов эукариот — прерывистость структуры смысловой части. Смысловые участки, несущие информацию о последовательности аминокислот в белке — экзоны, чередуются с участками некодирующих последовательностей — нитронами. Часто интроны по длине могут превосходить экзоны. Наличие избыточных последовательностей приводит к тому, что длина гена может быть в несколько раз больше, чем требуется для кодирования аминокислот в белке Гаплоидный набор9 хромосом человека содержит 3,5 х 10 нуклеотидных нар, что но количеству соответствует примерно 1,5 млн. пар генов. Однако данные по изучению генома человека показывают, что организм человека имеет не более 100 тыс. генов. Это значит, что в клетках человека только 1% ДНК выполняет кодирующие функции. В отношении оставшихся 99% существуют разные гипотезы, обосновывающие их регуляторные и структурные функции. Для человека данные об экзонноинтронном строении генов впервые были получены для гена гемоглобина, а затем и для многих других генов. Это открытие стало возможным в конце 70-х гг. благодаря разработке метода синтеза комплементарных ДНК на матрице м-РНК с помощью обратной транскриптазы, а также благодаря освоению целого ряда методов генно-инженерной техники. В них входит разрезание генома с помощью рестриктаз на последовательности, соответствующие отдельным генам, выделение таких генов, встраивание их в векторные молекулы, внедрение векторов в клетки бактерий и клонирование там таких рекомбинантных молекул, что позволяет получать банки генов.

Процесс транскрипции на ДНК, как на матрице, связан с синтезом комплементарной последовательности РНК, включающей и интроны, и экзоны. Затем в ходе созревания РНК в ядре из нее удаляются интроны, а концы соседних экзонов сшиваются стык в стык. Процесс удаления последовательностей РНК, соответствующих интронам, и соединение участков с транскрибируемыми последовательностями экзонов называется сплайсингом. Кроме того транскрибируемая молекула модифицируется добавлением метилированного Г-нуклеотида на 5'-конце (кэпирование) и поли-А последовательности на 3'-конце. Модифицированные участки играют важную роль в инициации белкового синтеза, защищают транскрипт т-РНК от деградации. Имеются данные, свидетельствующие о том, что поли-А конец участвует в транспорте зрелой м-РНК из ядра в цитоплазму и продлевает ее функционирование там. Весь суммарный процесс формирования зрелых молекул РНК из предшественников называется процессингом. Созревшая м-РНК выходит в цитоплазму, прикрепляется к рибосоме, где генетическая информация транслируется в белковую последовательность (рис. 3.15).
Большинство интронных последовательностей, по-видимому, не обладают специфическими функциями. Однако один и тот же транскрипт РНК может подвергаться сплайсингу по-разному, следовательно с одного транскрипта в ходе сплайсинга способны образоваться несколько различных РНК. Такой сплайсинг называется альтернативным. Альтернативный сплайсинг сообщает клетке дополнительную генетическую пластичность. Рассмотрим это явление на примере белка фибронектина.
В плазме крови человека присутствует белок фибронектин. Он синтезируется клетками печени и выделяется в кровь. Этот же белок может синтезироваться клетками соединительной ткани, эндотелиальными клетками, выстилающими кровеносные сосуды, и некоторыми другими типами клеток. В этом случае фибронектин в нерастворимой форме накапливается в межклеточном пространстве. Функции фибронектина многоплановы: он играет роль в поддержании гомеостаза, в процессах тромбоза, миграции и дифференцировки клеток сосудов, в развитии атеросклеротических бляшек в сосудах. Известно много вариантов фибронектина, он обладает ткане и возрастной специфичностью. Показано, что варианты фибронектина образуются с одного геномного транскрипта за счет альтернативного сплайсинга
Вариации м-РНК-последовательностей фибронектина связаны с присутствием или отсутствием по крайней мере двух интронов.
Известные вариации иммуноглобулинов также в значительной мере обеспечиваются процессом созревания новосинтезированных гигантских м-РНК в ядре.
В начале каждого гена, до его смысловой части, представленной экзонами, находятся участки, которые обеспечивают регуляцию работы гена. К числу регуляторных участков, одинаковых для всех генов, относятся ТАТА-последовательности (рис. 3.14), где чередуются тимин и аденин ("ТАТА-БОКС"). Этот участок лежит на 30 нуклеотидов левее места начала считывания гена. Установлено, что РНК-полимераза, фермент осуществляющий транскрипцию, так ложится на ДНК, что ее опознающая часть закрывает "TATA-БОКС", а ее активный центр оказывается над первым считываемым нуклеотидом. Далее по длине гена следует промоторный участок, который способствует правильной установке рамки считывания нуклеотидов, поскольку процесс считывания генетического кода не допускает возможности перекрывания кодонов
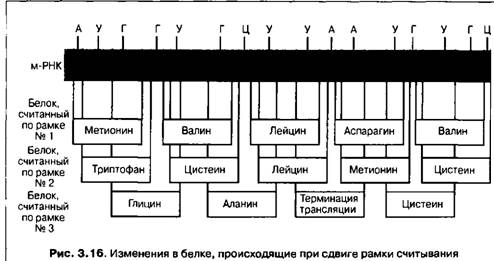
Иногда изменения рамки считывания могут происходить из-за выпадения или добавления одного или нескольких нуклеотидов, тогда при последующей сборке белка в нем будет нарушена последовательность аминокислот. Такая ситуация получила название мутации со сдвигом рамки (рис. 3.16).
За промоторным участком следует палиндром ("перевертыш"), или инвертированный повтор. Этот участок ДНК одинаково читается в обоих направлениях и имеет центральную точку, относительно которой последовательность остается одинаковой в обеих цепях ДНК. Следовательно, такой участок ДНК имеет две оси симметрии : вдоль и поперек. Важное свойство палиндромов — возможность образовывать шпильки в РНК или структуры креста в ДНК за счет комплементарного взаимодействия не между двумя нитями ДНК, а между нуклеотидами каждой цепи. В результате этого палиндром ДНК превращается в крест, что делает невозможным дальнейшее продвижение фермента РНК-полимеразы, и процесс транскрипции прекращается, если рамка считывания установлена неверно.
В последнее время описаны специфические регуляторы работы некоторых генов — энхансеры
Они расположены впереди гена на расстоянии в сотни и тысячи нуклеотидных нар от него. У эукариот существуют специальные регуляторные белки, опознающие энхансер и присоединяющиеся к нему, в результате чего происходит активация работы гена.
По данным разных авторов, содержание ДНК в диплоидной клетке человека составляет примерно 7,3 х 10~2г, что соответствует 7,1 х 109 нулеотидных пар. Каждая молекула ДНК гетерогенна но своему составу. В ней встречаются участки с уникальной последовательностью азотистых оснований, которые несут информацию для большинства белков клетки. В то же время в ней встречаются последовательности нуклеотидов, многократно повторяющиеся в геноме в составе этой же или других молекул ДНК. Такие повторяющиеся последовательности подразделяют на два класса. Первый — умеренно повторяющиеся последовательности с числом повторов от 102 до 105 на геном. На их долю приходится примерно четверть ДНК, и они представляют собой блоки истинных генов, как, например, гены гистонов (рис. 3.17). К этому же классу умеренных повторов относятся короткие последовательности, которые не кодируют белки, они разбросаны по всему геному, а их длина соответствует примерно 300 н п (нуклеотидных пар). Второй класс — часто повторяющиеся последовательности, или сателлитные ДНК, число повторов которых на геном превышает миллион (1х106) раз. Это нетранскрибирующиеся участки ДНК, на которых не идет образование РНК.
В работах разных авторов показано, что лишь немногим более 50% ДНК генома человека представлено уникальными фрагментами длиной около 2000 н.п., и далеко не все из них представляют структурные гены. В основном они распределены в составе длинных молекул ДНК между короткими, умеренно повторяющимися последовательностями, длина которых не превышает 300 н.п. Многие из таких умеренных повторов имеют сходное строение. Вероятно, они выполняют структурные и регуляторные функции в составе генов. Умеренные повторы, представляющие гены, встречаются в каждой клетке человека, где есть ядро. Они содержат гены, необходимые всем клеткам в каждой фазе индивидуального развития. Это гены рибосомной РНК, гистонов и транспортной РНК.
Гены рибосомной РНК являются частью района ядрышкового организатора на хромосомах. Они кодируют рибосомные РНК, которые синтезируются в ядрышке
В ядрышке происходит процессинг рибосомных РНК: из одной гигантской новосинтезированной молекулы образуются три разные молекулы рибосомных РНК меньшего размера, а избыточные последовательности удаляются. Зрелые рибосомные РНК участвуют в сборке субъединиц рибосом. Таким образом, ядрышко — это место в ядре, где функционируют рибосомные гены, оно содержит совокупность всех рибосомных РНК, находящихся на разных стадиях процессинга, здесь происходит процесс формирования субъединиц рибосом. У человека район ядрышкового организатора расположен в коротких плечах акроцентрических хромосом: 13-й, 14-й, 15-й, 21-й и 22-й (рис. 3.18). В формировании ядрышка могут принимать участие сразу несколько хромосом. Среднее число копий рибосомных генов в клетках человека по данным разных авторов 416-443 на диплоидный геном.


К классу умеренно повторяющихся последовательностей у человека можно отнести семейство многочисленных генов вариабельных участков иммуноглобулинов, играющих ключевую роль в иммунитете. Умеренные повторы обнаружены во всех хромосомах человека, где они локализованы по всей длине плеч
Часто повторяющиеся последовательности занимают примерно десятую часть генома. Участки ДНК с такими повторами выявлены в определенных местах хромосом: это около-центромерные и теломсрные районы. На дифференцированно окрашенных хромосомах они выявляются в виде гетерохроматина. Причем это гетерохроматин конститутивный — в нем не происходит транскрипция. Более того, конститутивный гетерохроматин может влиять на близко расположенные гены, подавляя их активность.
Частые повторы обычно не бывают длинными. Так, теломерные районы хромосом человека, ограничивающие хромосомы с двух концов, содержат 250-1500 повторов ТТАГГ. Эти повторы играют очень важную роль: предотвращают слипание хромосом концами и их укорачивание при многократных репликациях ДНК в связи с клеточными делениями. Часто повторяющиеся последовательности ДНК иначе называют сателлитной ДНК. Сателлитная ДНК не всегда имеет характер видового признака, как это видно на повторах теломерных районов. Однако у человека выделены и охарактеризованы индивидуальные сателлитные ДНК, расположенные в разных хромосомах. Так, известны несколько типов сателлитной ДНК из Y-хромосомы и 1-й, 9-й и 16-й хромосом
Функции сателлитной ДНК во многом остаются неизвестными. Предполагается, например, что сателлитная ДНК участвует в распознавании гомологичных хромосом во время конъюации в мейозе, рассматривается также возможность регуляторного участия сателлитной ДНК в функционировании генов.
Пришедшие в генетику новые методы позволили расширить знания о структуре генетического материала. По современным данным, он оказался намного менее статичен, чем представлялось раньше. Так, например, известны описанные Барбарой Мак-Клинток мобильные контролирующие генетические элементы в геноме кукурузы, способные перемещаться с одного гена на другой, увеличивая их нестабильность. Соматическими мутациями, связанными с присутствием мобильных контролирующих элементов, обусловлена мозаичная окраска початков у кукурузы. Найдены мобильные генетические элементы и у дрожжей. Позже было выявлено несколько классов мобильных генетических элементов у бактерий и показано, что они могут встраиваться во многие участки генома клетки хозяина. В зависимости от структуры мобильного генетического элемента внедрение бывает строго специфичным или случайным
Установлено, что при внедрении мобильного элемента встраивается не он сам, а его копия, в то время как исходный элемент остается на своем месте. Встраивание мобильного элемента в структурный ген приводит к мутации. Кроме того, перемещение мобильных элементов может стимулировать хромосомные аберрации. Для выявленных у дрозофилы мобильных генетических элементов показано, что они могут образовываться из рассеянных по геному генных элементов, из повторяющихся последовательностей конститутивного гетерохроматина прицентромерных районов хромосом, а также иметь вирусную природу. Все мобильные элементы обладают некоторыми общими чертами структурной организации — в частности инвертированными повторами на концах. Их репликация независима. Кроме того, они способны вызывать мутации с высокой частотой, причем мутации нестабильные, часто ревертирующие к исходному состоянию. Существуют предположения, что перенос генов мобильными элементами является одним из факторов эволюции. Например, последовательности ДНК, гомологичные глобиновому гену человека, были обнаружены у бобовых растений. Функция такой структуры может заключаться в обеспечении кислородом клубеньковых бактерий. А наличие такого гена в растениях может быть объяснено переносом его от насекомых или млекопитающих. Следует, однако, заметить, что у человека подобные элементы в геноме еще не выявлены. Тем не менее в геноме человека обнаружены повторяющиеся последовательности, содержащие палиндромы с инвертированными повторами, которые по
аналогии напоминают мобильные элементы. Первым этапом в транспозиции некоторой последовательности ДНК должно быть образование внехромосомных кольцевых копий этого участка. Подобные кольцевые структуры обнаружены в стареющих фибробластах в клеточных культурах человека. Кроме того для генома человека описано явление конверсии глобиновых генов, когда один аллель модифицируется другим, в результате чего гетерозигота становится гомозиготой.
Новые данные углубляют понимание структурной организации генетического материала и механизмов его работы. Однако по-прежнему верным остается постулат о стабильности генетического аппарата, на котором основаны все закономерности наследования признаков. Благодаря стабильности генома, существует наследственность, обеспечивающая преемственность фенотипических признаков из поколения в поколение.
– Конец работы –
Эта тема принадлежит разделу:
Тема ХРОМОСОМНАЯ ТЕОРИЯ
Хххххххх хххххххх хххххх Тема ХРОМОСОМНАЯ ТЕОРИЯ... Занятие Генные мутации... Занятие Хромосомные и геномные мутации...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Организация генетического материала в хромосомах человека
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.022 сек.
Новости и инфо для студентов