Соотношения Харди—Вайнберга доказывают генетическую основу групп крови системы АВО - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА Множественный Аллелизм. До Сих Пор Мы Рассматривали В Каждом Локусе То...
Множественный аллелизм. До сих пор мы рассматривали в каждом локусе только два разных аллеля. Однако часто встречаются ситуации когда локус может быть представлен более чем двумя аллелями. Примеры такого «множественного аллелизма» описаны и для людей, и для экспериментальных животных. Наиболее известны среди них - серия аллелей локуса white у Drosophila melanogaster и серия альбино-локуса у кроликов.
Важно помнить, что
а) любой индивид может иметь максимум два аллеля из серии (если у него две гомологические хромосомы, а, например, не три, как у трисомиков);
б) кроссинговер между этими аллелями может не учитываться, поскольку они расположены в гомологичных локусах. С уточнениями этого второго условия мы будем иметь дело в разд. 3.5 в связи с обсуждением современных концепций гена. Здесь же будет описана простейшая формальная модель на примере групп крови АВО.
Генетика групп крови АВО. Группы крови АВО были открыты в 1900 г. Ландштейнером [259].
| Таблица 3.4.Сравнение двух гипотез о наследовании групп крови АВО. (По Wiener A.S, 1943, с изменениями.)
|
| Генотипы родителей
| Генотипы детей, ожидаемые в соответствии с гипотезой
|
| двух разных диаллельных генах
| множественных аллелях одного локуса
|
| 0 x 0
|
|
|
| 0 х А
| 0,А
| 0,А
|
| 0 х В
| 0,В
| 0,В
|
| А х А
| 0,А
| 0,А
|
| А х В
| 0,А,В,АВ
| 0,А,В,АВ
|
| В х В
| 0,В
| 0,В
|
| 0 х АВ
| 0,А,В,АВ
| А,В
|
| А х АВ
| 0,А,В,АВ
| А,В,АВ
|
| В х АВ
| 0,А,В,АВ
| А,В,АВ
|
| АВ х АВ
| 0,А,В,АВ
| А,В,АВ
|
Несчастные случаи, имевшие место при переливании крови, заставили задуматься над причиной и тем самым способствовали открытию системы АВО. Первая разумная генетическая теория предложена фон Дунгерном и Гиршфельдом (1911) [631]. Для объяснения четырех фенотипов А, В, О и АВ они предположили наличие двух независимых пар генов (А,О; В,О), где А и В - доминантные аллели. Бернштейн (1925) [574] проверил эту гипотезу, используя прежде всего соотношения, ожидаемые из закона Харди-Вайнберга. Он пришел к выводу об ошибочности их концепции и предложил правильное объяснение - три аллеля с шестью генотипами и четырьмя фенотипами вследствие доминирования аллелей А и В над аллелем О.
Наиболее очевидный способ проверки этих гипотез - обратиться к семейным исследованиям. Однако различия между этими двумя гипотезами ожидаются только для тех браков, в которых по крайней мере один из родителей имеет группу АВ (табл. 3.4): двухлокусная диаллельная модель допускает в таких браках наличие детей с группой О, а трехаллельная монолокуснаянет. Хотя группа АВ самая редкая, в литературе уже публиковались сообщения о детях с группой крови О у родителей с группой АВ (либо группа крови в таких случаях неправильно диагностирована, либо дети внебрачные). Однако эти наблюдения не ввели Бернштейна в заблуждение. Его рассуждения были следующими. Предположим, что справедлива теория двух генов. Пусть р будет частотой аллеля А (1 — р = р' – частота
3. Формальная генетика человека 177
аллеля а) и q - частотой аллеля В (1 — q = q' – частота аллеля b). Тогда в популяции следует ожидать такие относительные частоты генотипов и фенотипов:
| Фенотип
| Генотип
| Частота
|
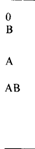
| 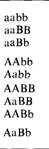
| 
|
| Это ведет к таким соотношениям (А, В – частоты фенотипов):
|

|
Различия настолько велики (и так убедительны), что их невозможно объяснить случайными отклонениями. Первой альтернативой, рассмотренной Бернштейном, было предположение о гетерогенности изучаемой популяции. Однако это объяснение оказалось недостаточным. С другой стороны, легко показать, что генотипические распределения во всех популяциях, для которых имелись данные, находились в прекрасном согласии с соотношениями, ожидаемыми на основе мультиаллельной монолокусной гипотезы.
Чтобы понять рассуждения Бернштейна, следует по-новому взглянуть на закон Харди-Вайнберга. Он был выведен нами для специального случая двух аллелей. Однако можно показать, что этот закон применим и ко многим аллелям. Если обозначить частоты п аллелей как р1 , р2 , ... рn, то относительные частоты генотипов задаются разложением (p1 + p2 + … + pn)2. Для конкретного случая аллелей А, В, и О с частотами p,q и r отсюда следует, что распределение генотипов имеет вид

|
| Для четырех классов фенотипов
|

|
| можно получить следующие вероятности:
|

|
| Отсюда следует
|

|
| поэтому
|

|
| и справедливо соотношение
|

|
Это равенство можно проверить, используя распределения фенотипов АВО в различных популяциях мира. Критерием служит равенство единице суммы аллельных частот, вычисленных по приведенной выше формуле. Кроме того, на основе получаемых частот аллелей можно вычислить ожидаемые частоты генотипов и сравнить их с наблюдаемыми. Помимо корректности самой генетической гипотезы этот результат подлежит проверке на выполнение другого условия: в популяции по изучаемому признаку должно быть случайное скрещивание (панмиксия).
Для данных, анализированных Бернштейном, уже было получено превосходное соотношение, и это подтвердилось для огромного количества накопленных с тех пор материалов. Для понимания принципа вычисления приведем один пример. В Берлине были найдены следующие фенотипические частоты групп крови.
| Все лица (n)
| А
| В
|
| АВ
|
| 21 104
| 9 123
| 2 987
| 7 725
| 1 269
|
|
| (43,23%)
| (14,15%)
| (36,60%)
| (6,01%)
|
178 3. Формальная генетика человека
По формуле Бернштейна получаем частоты аллелей:

Итак, р + q + r = 0,9991.
На первый взгляд этот результат хорошо согласуется с ожидаемым значением 1. В качестве статистического критического критерия для тестирования значимости отклонения можно использовать метод хи-квадрат [625]:

В нашем примере χ12 = 0,88, что явно меньше граничного значения χ12 = 3,84 при одной степени свободы (для 5%-ного уровня значимости). Следовательно, подтверждается, что найденные значения хорошо согласуются с генетической гипотезой и с предположением о случайном скрещивании для системы АВО.
Позже Бернштейн показал, каким образом разность D можно использовать для коррекции вычисленных частот аллелей. Если обозначить непоправленные частоты р', q' и r', то можно применить следующие формулы:
p=p'(1+D/2),
q = q'(1+D/2),
r = (r' + D/2)(l + D/2),
что в нашем случае дает
р = 0,2876(1 + 0,00045) = 0,2877,
q = 0,1065(1 + 0,00045) = 0,1065,
r = (0,6050 + 0,00045)(1 + 0,00045) = 0,6057.
В процессе тестирования двух гипотез о наследовании групп крови АВО Бернштейн разработал методы вычисления частот аллелей. Эти методы приобрели практическое значение и будут подробно рассмотрены нами отдельно (приложение!).
Значение равновесия Харди-Вайнберга. Если наблюдаемые доли генотипов в популяции соответствуют ожидаемым из закона Харди-Вайнберга, то говорят, что популяция находится в равновесии Харди-Вайнберга. Такое равновесие следует отличать от равновесия между аллелями, которое будет рассматриваться при обсуждении отбора (разд. 6.2.1) и мутаций (разд. 5.1.3). Равновесие Харди-Вайнберга - это равновесие распределения генов в популяции (пула генов) между разными генотипами. Если равновесие было нарушено какими-либо силами, то при случайном скрещивании оно восстановится за одно поколение. Следует помнить, однако, что закон Харди-Вайнберга справедлив при следующих условиях.
1. Скрещивания в отношении изучаемых фенотипов должны быть случайными. Это условие можно уверенно считать выполняющимся для таких признаков, как группа крови или полиморфные ферменты. Но вряд ли оно справедливо для морфологических признаков, таких, как рост, а тем более для поведенческих, таких, как интеллект. Об этом необходимо помнить, когда используемые в количественной генетике меры сходства, например корреляции между родственниками, интерпретируются в генетических терминах.
2. Отклонение от случайного скрещивания может быть вызвано, в частности, кровнородственными браками: если в популяции высокий уровень кровного родства, то следует ожидать увеличения количества гомозигот (разд. 6.3.1). Можно даже вычислить частоту кровнородственных браков в популяции на основе отклонений от соотношений Харди-Вайнберга.
3. Соотношения Харди-Вайнберга могут быть нарушены недавними миграциями.
4. Иногда в качестве фактора, нарушающего соотношение Харди-Вайнберга, упоминается отбор. Это справедливо, но не обязательно. Как правило, отбор изменяет генные частоты. Однако отбор, действующий до репродуктивного возраста, например в пренатальном периоде или позже, включая детство и юность, совсем не влияет на соотношения Харди-Вайнберга у взрослых. Отклонения могут наблюдаться в субпопуляции детей в зависимости от конкретного типа отбора. Кроме того, даже при сильном отборе в соответствующей возрастной группе регистрация статистически значимых отклонений от соотношений Харди-Вайнберга требует выборок большого объема - больше, чем обычно имеется в распоряжении исследователя. Иногда вывод об отсутствии значимого отбора фор-
3. Формальная генетика человека 179
мулируется исходя из наблюдаемого соответствия популяции соотношениям Харди-Вайнберга. Однако такой вывод, если он тщательно не проверен, легко может оказаться ошибочным. Рассматривая все теоретические возможности нарушения соотношений, поражаешься тому, как часто в человеческих популяциях, выполняется соотношение Харди-Вайнберга.
5. Формально отклонение от закона Харди-Вайнберга может наблюдаться в том случае, если популяция представляет собой смесь субпопуляций, лишь частично скрещивающихся между собой (случайное скрещивание происходит только внутри субпопуляций, и, следовательно, генные частоты в этих субпопуляциях различны). Впервые на это указал Валунд (1928) [929], который вывел формулу для вычисления коэффициента инбридинга F на основе дисперсий генных частот между субпопуляциями.
6. Другой причиной отклонения может служить существование пока не выявленного («немого») аллеля, в связи с чем гетерозиготные носители этого аллеля неотличимы от гомозигот по обычному аллелю. Однако Смит (1970) [879] указал, что генетически немой аллель может вызывать значимое отклонение от закона Харди-Вайнберга только тогда, когда гомозиготы по этому аллелю имеют частоту, достаточно высокую, чтобы быть выявленными.
Все темы данного раздела:
Фогель Ф., Мотульски А.
Ф74 Генетика человека: В 3-х т. Т. 1: Пер. с англ.-М.: Мир, 1989.-312 с, ил.
ISBN 5-03-000287-1
Книга двух известных генетиков из ФРГ и США является фундаментальным учебником по г
История генетики человека
История генетики человека представляет особый интерес, поскольку концепции этой науки часто оказывали влияние на социальные и политические события. В то же время развитие генетики ч
Ученые до Менделя и Гальтона
Литература средневековья содержит не много упоминаний о наследственности.
Анализ природных явлений привел к созданию современной науки и возникновению нового взгляда на человека. Эмпиричес
Прикладные исследования применительно к человеку: «врожденные ошибки метаболизма» по Гэрроду
Вэтом историографическом введении будет описан только первый шаг в развитии таких исследований: статья Гэррода (Garrod, 1902) [249] «Распространенность алкаптонурии: изучение химических особенносте
Видимые носители генетической информации: первые исследования хромосом
И биометрический анализ Гальтона, и опыты Менделя по скрещиванию основывались
1 Доклад Комитету по вопросам эволюции Королевского общества (1902 г.)
&nb
Группы крови АВО
АВО-система групп крови была открыта Ландштейнером в 1900 г. [259]. В 1911 г. ван Дунгерн и Гиршфельд [245] подтвердили, что группы крови наследуются. Эти факты доказали применимость законов Мендел
Закон Харди-Вайнберга
Английский математик Харди [252] и немецкий врач Вайнберг [289] примерно одновременно в (1908 г.) доказали основополагающую теорему популяционной генетики, которая объясняет, почему от поколения к
Достижения генетики человека в период 1910-1930 гг.
В период 1910-1930 гг. в генетике человека не было сделано новых фундаментальных открытий. Основная часть представлений классической генетики (таких, как сцепление, нерасхождение, скорость мутацион
Генетика человека, евгеника и политика
1.8.1. Великобритания и США [236; 246; 256; 263; 283]
Первое десятилетие нашего века ознаменовалось развитием евгеники в Европе и Соединенных Штатах Америки. На многих уче
Генетика поведения человека
Как на Востоке, так и на Западе продолжаются ожесточенные дискуссии о роли генетических факторов в детерминации поведения человека, его интеллекта и личностных особенностей. Некоторые ученые отрица
Генетическая эпидемиология
В 40-50-е гг. существовало несколько институтов, занимавшихся пионерскими исследованиями в области эпидемиологии генетических заболеваний. Институт Кемпа в Копенгагене, отделы Нила в Энн Арборе, шт
Биохимические методы
После второй мировой войны благодаря появлению биохимических и цитологических методов произошло быстрое возрождение генетики человека. Генетика человека, которой в основном занимались ученые, испол
Индивидуальные биохимические различия
Изучение вариантов фермента глюкозо-6фосфат-дегидрогеназы (G6PD) помогло развить представления о значительной мутационной изменчивости. Наличие индивидуальных биохимических особенностей объясняет р
Цитогенетика, генетика соматических клеток, пренатальная диагностика
Совершенствование цитогенетических методов сделало возможным их применение для изучения многих типов врожденных аномалий и интерсексов. Было показано, что возникновение специфической формы рака, хр
Методы исследования ДНК в медицинской генетике
Достижения молекулярной генетики и развитие методов исследования ДНК быстро нашли применение для решения практических задач медицинской генетики. Поскольку наиболее существенные успехи в изучении г
Нерешенные проблемы
Генетика человека большинством своих достижений обязана тому, что она опиралась на законы Менделя и использовала методы, разработанные в различных областях биологии. Такие важные проблемы, как регу
Цитогенетика человека – запоздалое, но счастливое рождение
Хромосомная теория менделевского наследования была сформулирована в 1902 г. Саттоном и Бовери. В том же году Гэррод, установив аутосомно-рецессивный тип наследования алкаптонурии и обсуждая в связи
История развития цитогенетики человека
Первые наблюдения митотических хромосом человека [522]. Можно сказать, что исследования по цитогенетике человека начались с работ Арнольда (1879) [297] и Флемминга (1882) [348], которые впер
Нормальный кариотип человека в метафазе митоза
Стандартное окрашивание. Хромосомы располагаются и нумеруются в зависимости от их длины. Согласно Денверской классификации, предложено нумеровать пары хромосом от 1 до 23 (1960). Патау (1960
Синдромы, связанные с аномалиями числа хромосом
Механизмы, лежащие в основе геномных мутаций (аномалии числа хромосом). Аномалии числа хромосом могут быть вызваны разными причинами: 1. Наиболее важным механизмом является нерасхождение. Хр
Кариотипы u клинические синдромы
Первые наблюдения синдрома Дауна. Как только трисомия 21 была идентифицирована как причина синдрома Дауна, естественно возник вопрос о том, у всех ли больных имеется эта трисомия. Если не у
Сегрегация и пренатальная селекция транслокаций: методологические аспекты
Проблема сегрегации и пренатальной селекции транслокаций служила предметом многих исследований, но результаты их оказались противоречивыми и в конечном счете не позволили дать общую картину. Совсем
Первые наблюдения
Нерасхождение половых хромосом и определение пола у дрозофилы. В 1910 г. Морган [448] впервые детально описал Х-сцепленное наследование и X-Y-тип хромосомного определения пола у Drosophil
Х-хромосо иные анеуплоидии у человека: современное состояние проблемы
Различие между Х-хромосомными и аутосомными анеуплоидиями. Вскоре после первых открытий анеуплоидии по половым хромосомам последовали и друше. Если рассматривать все эти случаи в целом как г
Организация генетического материала в хромосомах человека
Два первых десятилетия современного этапа в изучении хромосом человека прояснили многие аспекты организации генетического материала. Однако мало было конкретной информации о том, как эти знания мог
Уникальная и повторяющаяся ДНК
Избыточность ДНК в геноме человека. Вскоре после того, как генетический код был расшифрован (в начале 60-х гг.), ученые пришли к выводу об избыточности ДНК в эукариотических клетках. По данн
Гетерохроматин
Определение и свойства. Термин «гетерохроматин» был предложен Хейтцем (Heitz, 1928) [469]. Он писал: «У P. (Pellia)epiphylla (мох) некоторые участки пяти из девяти хромосом ведут себя
Интеграция хроматиновых волокон в хромосомную структуру
Интерфаза. Интерфазная хромосома представляет собой элементарную фибриллу, состоящую из нуклеосом, соединенных линкерами. Эта фибрилла пронизывает не все ядро, а лишь определенные его област
Интегральная модель структуры хромосомы
Эти данные вместе с результатами молекулярно-биологических исследований (см. ниже) позволяют сформулировать интегральную модель хромосомы: она состоит из единственной двойной спирали ДНК, объединен
Генетический код
Одним из важнейших достижений 60-х гг., ознаменовавшим возникновение новой генетики, было открытие генетического кода. Благодаря использованию синтетических тринуклеотидов удалось показать, что каж
Анализ гена человека
β-глобиновый ген. Молекула гемоглобина, а также клинические и биохимические последствия ее изменений будут детально описаны в разделе 4.3. Гемоглобин взрослого человека НbАХ
Рестрикционные эндонуклеазы
Первые наблюдения. Заражая фагом X различные штаммы Е. coli, Арбер [296] обнаружил, что ДНК этого фага при пассаже через бактерию разрезается и теряет свою инфекционность. Оказ
Гибридизация нуклеиновых кислот
Принцип. В разд. 2.3.1.1 мы уже упоминали метод идентификации ДНК-повторов, основанный на разделении двойных цепей при повышении температуры и их реассоциации при быстром снижении температур
Сортировка хромосом при помощи цитофлуорометрии
Зачем нужны сортировка хромосом и препараты отдельных хромосом? Сортировка хромосом методом цитофлуорометрии используется в двух разных целях: 1) для идентификации и количественного анализа
Анализ β-глобинового гена и обобщение опыта исследования одного гена.
β-глобиновый ген. Разд. 2.3.2.1 начинался с анализа структуры β-глобинового гена, которая была раскрыта благодаря использованию новых методов и подходов. Наиболее важные из них опи
Структура гена фактора VIII (антигемофилический фактор)
Антигемофшшческий фактор (фактор VIII). Гемофилия А — «классическая» наследственная болезнь с Х-сцепленным типом наследования (разд. 3.1.4). Анализ процесса свертывания крови позволил в 50-х
Семейства генов
Примеры семейства генов. Под семейством генов мы понимаем группу функционально родственных генов, имеющих сходную структуру и общее происхождение. Ярким примером генного семейства являются д
Динамичность генома
Методы новой генетики расширили наши знания о структуре генетического материала. Представление о хромосоме как о нитке с бусинами-генами соответствует реальным фактам теперь еще в меньшей степени,
Геном митохондрий
Структура и функция митохондрий. Митохондрии - это цитоплазматические органеллы. Их количество и форма варьируют в зависимости от функции клетки. Например, у млекопитающих в клетках печени и
Новая генетика и концепция гена
Молекулярная цumoгенетика. Методы новой генетики важны не только для изучения структуры генов, они оказывают все возрастающее влияние на эффективность цитогенетической диагностики. Например,
Кодоминантный тип наследования
Первые случаи кодоминирования у человека были обнаружены при изучении наследования групп крови, например, системы MN (табл. 3.1). С развитием методов генетического анализа на уровне белков было отк
Аутосомно-доминантиый тип наследования
Первое описание родословной с аутосомно-доминантным наследованием аномалии у человека дано в 1905 г. Фараби [656]. В учебниках это заболевание обычно называют брахидактилией (короткопалость), но из
Аутосомно-рецессивный тип наследования
Тип наследования называется рецессивным, когда гетерозигота фенотипически не отличается от нормальной гомозиготы. Однако во многих случаях с помощью специальных методов между ними можно обнаружить
Х-сцепленные типы наследования
У человека каждый брак можно рассматривать как менделевский бэккросс (возвратное скрещивание) в отношении Х- и Y-xpoмосомы:
Отцовские гам
Родословные, не соответствующие простым типам наследования
Время от времени публикуются сообщения о родословных, которые трудно согласовать с каким-либо простым типом наследования. Нередко причины этого связаны с ошибками регистрации или документации. Но е
Гены-модификаторы
До сих пор мы рассматривали только моногенно контролируемые признаки. Однако на фенотипическое проявление одного гена обычно влияют другие гены. Эксперименты на животных, особенно на млекопитающих,
Количество известных заболеваний человека с простым типом наследования
Многие годы Мак-Кьюсик [133] собирает и документирует заболевания человека с простым типом наследования. Табл. 3.3 основана на шестом издании его книги с более поздними дополнениями. Со времени опу
Формулировка и вывод закона
До сих пор применение законов Менделя к наследованию признаков у человека мы рассматривали с точки зрения изучения отдельных семей. Каковы, однако, следствия из этих законов для генетической структ
Генные частоты
Одна пара генов — два фенотипа. Редкие аутосомно-рецессивные заболевания контролируются только одной парой генов, и обычно известны только два фенотипа: гетерозиготы либо не идентифицируются
Сегрегационные отношения как вероятности
В ходе мейоза при отсутствии каких-либо нарушений различные гаметы образуются в точно таких относительных пропорциях, какие ожидаются из законов Менделя. Из диплоидного гетерозиготного сперматоцита
Простые вероятностные проблемы в генетике человека
Независимые события и прогноз при медико-генетическом консультировании. Врач, давший ошибочный прогноз супружеской паре с двумя детьми-альбиносами, не принял в расчет, что зачатия каждого из
Тестирование сегрегационных отношений в отсутствие смещений, связанных с регистрацией: ко доминантное наследование
За исключением предельных случаев, вычисление точных вероятностей семей или групп семей определенного типа обычно не практикуется. Следовательно, применяемые статистические методы либо основываются
Тестирование сегрегационных отношений: редкие признаки
Основные типы смещений. Если изучаемый признак редкий, то обычно семьи не регистрируют случайно, а начинают с «пробанда», т. е. индивида с данным признаком. Это приводит к смещениям вслед
Заболевания со сложным типом наследования
Обсуждавшиеся до сих пор методы применялись в основном при анализе признаков, наследующихся в соответствии с простыми менделевскими правилами. Однако для многих заболеваний, особенно для широко рас
Сцепление: локализация генов на хромосомах
Гены расположены в хромосомах в линейном порядке. Отсюда следует, что, во-первых, гены, локализованные в одной хромосоме, передаются совместно, а во-вторых, сегрегация сцепленных генов не является
Анализ сцепления у человека: гибридизация клеток и ДНК-технология
Слияние клеток: первые наблюдения. История открытия этого феномена описана Харрисом [702; 692]. Еще в 1838 г. Мюллер наблюдал двухъядерные клетки в опухолях, впоследствии Робин обнаружил их
Некоторые особенности генетической карты человека
Типы генных кластеров. При поверхностном знакомстве с генетической картой человека может возникнуть впечатление, что большинство локусов распределены в значительной мере случайно. Однако име
Почему существуют кластеры генов?
Генные кластеры - результат эволюционного процесса. В некоторых случаях кластеризация генов отражает историю эволюционного развития. Допустим, на ранних этапах эволюции существовал один локу
Группы крови: Rh-комплекс, неравновесие по сцеплению
История. В 1939 г. Левин и Стетсон [762] исследовали сыворотку крови женщины, которая родила мертвый плод и в анамнезе которой имело место переливание крови мужа, совместимой по АВО группе.
Генетическая детерминация мимикрии у бабочек
Отметим сразу, что знакомство с этим разделом не обязательно для понимания принципов генетики человека.
Ложная предупредительная окраска [866]. В процессе эволюции у некоторых живот
Гены Х-хромосомы человека, имеющие родственные функции
В ходе эволюции млекопитающих Х-хромосома в отличие от аутосом оставалась относительно неизменной. Существует несколько фактов, указывающих на гомологию Х-хромосом разных видов [156]. Важная особен
Неравный кроссинговер
Открытие неравного кроссинговера. В первые годы работы с дрозофилой некоторые авторы обратили внимание на то, что мутация Bar (Х-сцепленный доминантный признак) иногда ревертировала к
Уровни генетического анализа
Теория наследственности, созданная Менделем на основании опытов по скрещиванию гороха (разд. 1.4), претерпела в своем развитии несколько этапов. По современным представлениям ген - это фрагмент дву
Генный уровень
Конечная цель генетического анализа - выявить различия на уровне ДНК, т. е. идентифицировать мутантный сайт. Последовательность нуклеотидов в ДНК содержит информацию для последовательности аминокис
Анализ продукта гена: биохимический уровень
В этом случае идентифицировать мутантный сайт внутри гена невозможно, можно только идентифицировать ген, в котором произошла мутация. Для этого существует несколько способов.
1. Специфичес
Качественный феногенетический анализ: простые типы наследования
В данном случае выводы основаны на анализе фенотипических различий, отдаленных от первичного действия генов. Тем не менее иногда соответствие генотипа и фенотипа
232 3. Формальная ген
Генетический анализ на уровне количественного фенотипа – биометрический уровень
Аддитивная модель. Во многих случаях фенотипическая изменчивость настолько сложна, что эффекты отдельных мутаций уже нельзя идентифицировать и приходится мириться с генетическими выводами (о
Концепция наследуемости
Концепция наследуемости широко применяется в количественной генетике. Градации изучаемого признака, выраженные в метрических единицах, могут быть названы «значениями». Значение, измеренное у
Количественная генетика; концепции Менделя и Гальтона
Как связаны между собой две концепции, на которых основывается генетика человека? Представление о гене возникло на основе экспериментов Менделя (разд. 1.4), концепция Гальтона опирается на корреляц
Описание модели: эксперименты на животных
В предшествующем разделе генетический анализ количественного признака на биометрическом уровне обсуждался в отношении нормальных признаков с унимодальным и почти нормальным распределением в популяц
Простая теоретическая модель
Напомним модель, описанную в разд. 3.6.1: две пары аллелей с равными и аддитивными вкладами и частотами р1 = р2 = q1 = q2
Качественные (или полуколичественные) критерии мультифакториального наследования.
Можно сформулировать четыре таких критерия.
1. Близнецовый критерий: если конкордантность монозиготных (МЗ) близнецов вчетверо выше, чем конкордантность дизиготных (ДЗ) близнецов, то мульт
Какой вывод следует сделать, если статистический анализ не дает четкого ответа?
Выше уже указывалось, что совместимость полученных данных с генетической моделью еще не означает, что эта модель истинная. Совершенно разные модели могут одинаково хорошо соответствовать од-
Индуцированные радиацией доминантные мутации у мыши: мутации главных генов, не выявленные у человека
Экспериментальная работа с млекопитающими, физиология развития которых ближе всего к человеку, показывает, как действие главного гена может быть скрыто за фенотипической изменчивостью организма. Та
Как анализировать мулыпифакториальный признак, если отдельные формы с простыми типами наследования выделить нельзя?
Сложный функциональный дефект вызывается комбинацией малых нарушений. Как упоминалось выше, аддитивно-полигенная модель, используемая для анализа мультифакториального наследования, является слишком
Новая стратегия исследований
Чтобы глубже проникнуть в механизмы мультифакториального наследования, необходимо изменить стратегию исследований. Если прямой путь от фенотипа к генотипу не дает положительных результатов,
Система АВО
Вскоре после открытия групп крови АВО были высказаны предположения об ассоциации этих антигенов с определенными заболеваниями. Первый этап исследований подобного рода достиг своей кульминации в 20-
Kell-система
Мутации системы Kell, акантоцитоз и хронический грануломатоз. Помимо ассоциаций ряда заболеваний с распространенными группами крови известны некоторые примеры наследственных аномалий, связан
Концепция: природа - воспитание. Близнецовый метод
При обсуждении методов количественной генетики часто ссылаются на близнецовые данные, используемые для количественной оценки степени генетической детерминации отдельных признаков. Действительно, бл
Исторические замечания
Открытие близнецового метода обычно приписывают Гальтону (1876) [675], который, следуя Шекспиру (осознанно или неосознанно), ввел альтернативные понятия «природа» и «воспитание». (В пьесе «Буря» Пр
Исходная концепция
В основе близнецового метода лежит тот факт, что МЗ близнецы развиваются из одной зиготы. Отсюда следует, что генетически они должны быть идентичными. (Группу генетически идентичных особей называют
Биология близнецовости
Дизиготные близнецы. Большинство млекопитающих (грызуны, хищники, некоторые копытные) имеет многочисленный помет. Во время овуляции яичники выделяют одновременно несколько яйцеклеток, каждая
Ограничения близнецового метода
Систематические различия между близнецами и неблизнецами. Цель близнецовых исследований заключается в получении результатов, применимых не только к близнецам, но и ко всей популяции. В любом
Применение близнецового метода для анализа альтернативных признаков
В этой области близнецовый метод может служить достижению трех целей.
1. Различия в конкордантности между МЗ и ДЗ близнецами можно использовать для оценки значимости генетической изменчиво
Близнецовые исследования других широко распространенных заболеваний
В табл. 3.31 приведен перечень заболеваний, для которых с помощью близнецового
Таблица 3.31.Выборки близнецов с мультифакториальными (исключая психические) заболеваниями.
Близнецовый метод в изучении признаков с непрерывным распределением
В какой степени изменчивость признака в популяции детерминирована генетическими факторами? Чтобы ответить на этот вопрос, признак нужно уметь измерить. Это может показаться самоочевидным, однако в
Значения оценок наследуемости: данные по росту
Высокая наследуемость показана для роста. Это означает, что изменчивость условий среды, в которой обитает популяция изучаемых близнецов, оказывает
3. Формальная генетика человека 289
Биологические и патофизиологические подходы к генетической этиологии широко распространенных заболеваний
Анализ гетерогенности. Идентификация моногенных форм. Часто возникает необходимость обосновать с помощью соответствующих клинических, лабораторных и генетических методов выделение в пределах







Новости и инфо для студентов