рефераты конспекты курсовые дипломные лекции шпоры

- Раздел История
- /
- Изменение энергетического обмена в ходе эволюции
Реферат Курсовая Конспект
Изменение энергетического обмена в ходе эволюции
Изменение энергетического обмена в ходе эволюции - раздел История, А.И.Зотин, А.А.Зотин НАПРАВЛЕНИЕ, СКОРОСТЬ И МЕХАНИЗМЫ ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ Как Отмечалось Выше, Вывод Дольника (1968), Согласно Которому По Мере Усложне...
Как отмечалось выше, вывод Дольника (1968), согласно которому по мере усложнения организации животных стандартный обмен заметно возрастает, подтверждается всеми известными нам данными. Чтобы сделать этот вывод еще более наглядным мы собрали данные, представленные в табл.6-11,13,15-24,26-37,39,40 и в формулах (87)-(94), (96-108), (110-128), в одном месте (табл.41).
Таблица41
Сводная таблица коэффициентов а (мВт) и критериев упорядоченности Сr в разных типах, классах и отрядах животных. N - число изученных видов, n - число измерений.
| Таксон | а | N | n | Сr |
| Простейшие |
| Тип Apicomplexa | 0.046 | 0.66 | ||
| Тип Sarcomastigophora | 0.081 | 1.16 | ||
| Тип Ciliophora | 0.313 | 4.48 | ||
| Низшие беспозвоночные | ||||
| Тип Ctenophora | 0.113 | 2.17 | ||
| Тип Coelenterata | 0.297 | 5.07 | ||
| Класс Scyphozoa | 0.179 | 3.48 | ||
| Класс Hydrozoa | 0.194 | 3.54 | ||
| Класс Anthozoa | 0.426 | 6.81 | ||
| Черви | ||||
| Тип Plathelminthes | 0.582 | 8.67 | ||
| Класс Cestoda | 0.249 | 3.71 | ||
| Класс Trematoda | 0.285 | 4.40 | ||
| Класс Turbellaria | 0.466 | 6.93 | ||
| Высшие беспозвоночные | ||||
| Тип Echinodermata | 0.356 | 7.25 | ||
| Класс Echinoidea | 0.249 | 5.37 | ||
| Класс Asteroidea | 0.310 | 6.53 | ||
| Класс Holothuroidea | 0.367 | 7.25 | ||
| Класс Ophiuroidea | 0.409 | 8.17 | ||
| Класс Crinoidea | 0.677 | 13.98 | ||
| Тип Arthropoda | 0.881 | 14.38 | ||
| Тип Arthropoda (без Insecta) | 0.653 | 9.70 | ||
| Класс Myriapoda | 0.405 | 5.78 | ||
| Класс Arachnida | 0.530 | 9.64 | ||
| Отряд Amblypygi | 0.228 | 5.72 | ||
| Отряд Acariformes | 0.277 | 4.87 | ||
| Отряд Scorpiones | 0.272 | 6.84 | ||
| Отряд Parasitiformes | 0.513 | 8.82 | ||
| Отряд Aranei | 0.601 | 11.19 | ||
| Отряд Solifugae | 0.746 | 17.39 | ||
| Отряд Opiliones | 3.019 | 48.51 |
| Класс Crustacea | 1.208 | 20.72 | ||
| Отряд Cirripeda | 0.191 | 3.12 | ||
| Отряд Cumacea | 0.318 | 6.89 | ||
| Отряд Mysidacea | 0.743 | 13.32 | ||
| Отряд Decapoda | 0.770 | 14.61 | ||
| Отряд Spelaogriphacea | 1.047 | 16.90 | ||
| Отряд Isopoda | 1.089 | 18.49 | ||
| Отряд Tanaidacea | 0.911 | 19.57 | ||
| Отряд Amphipoda | 1.362 | 21.46 | ||
| Отряд Copepoda | 1.292 | 22.17 | ||
| Отряд Phyllopoda | 1.769 | 30.15 | ||
| Отряд Ostracoda | 1.129 | 30.26 | ||
| Отряд Euphasiacea | 1.976 | 32.51 | ||
| Отряд Anostraca | 5.955 | 122.39 | ||
| Класс Insecta | 1.792 | 33.10 | ||
| Отряд Collembola | 0.630 | 8.32 | ||
| Отряд Blattodea | 0.995 | 16.42 | ||
| Отряд Hemiptera | 1.238 | 19.43 | ||
| Отряд Dermoptera | 1.328 | 19.78 | ||
| Отряд Isoptera | 1.197 | 29.14 | ||
| Отряд Plecoptera | 2.024 | 30.15 | ||
| Отряд Orthoptera | 2.118 | 34.40 | ||
| Отряд Homoptera | 2.046 | 36.64 | ||
| Отряд Odonata | 1.957 | 44.32 | ||
| Отряд Coleoptera | 1.235 | 26.08 | ||
| Отряд Lepidoptera | 2.096 | 44.09 | ||
| Отряд Diptera | 2.894 | 48.43 | ||
| Отряд Hymenoptera | 1.330 | 24.31 | ||
| Сем. Apidae | 4.122 | 73.47 | ||
| Сем. Formicidae | 0.654 | 12.59 | ||
| Тип Mollusca | 1.060 | 21.03 | ||
| Тип Mollusca (без Cephalopoda) | 0.670 | 12.10 |
| Класс Bivalvia | 0.603 | 10.64 | ||
| Отряд Taxodonta | 0.45 | 6.94 | ||
| Отряд Eulamellibranchia | 0.54 | 8.41 | ||
| Отряд Dysodonta | 0.76 | 12.94 | ||
| Класс Gastropoda | 0.689 | 13.86 | ||
| Отряд Gymnosomata | 0.687 | 10.40 | ||
| Отряд Basommatophora | 0.616 | 11.57 | ||
| Отряд Archaegastropoda | 0.571 | 12.29 | ||
| Отряд Mesogastropoda | 0.650 | 13.35 | ||
| Отряд Nudibranchia | 1.154 | 17.21 | ||
| Отряд Stylommatophora | 0.982 | 18.07 | ||
| Отряд Neogastropoda | 0.817 | 18.09 | ||
| Отряд Sacoglossa | 0.980 | 25.17 | ||
| Отряд Cephalaspiea | 1.118 | 29.95 | ||
| Класс Loricata | 0.802 | 16.45 | ||
| Класс Cephalopoda | 4.46 | 99.77 | ||
| Отряд Nautilida | 1.03 | 27.6 | ||
| Отряд Octopoda | 1.87 | 50.08 | ||
| Отряд Decapoda | 6.63 | 142.57 | ||
| Тип Chaetognatha | 0.351 | 9.52 | ||
| Тип Tunicata | 0.200 | 5.29 | ||
| Тип Vertebrata | 6.93 | 33.61 | ||
| Водные и полуводные | ||||
| Класс Amphioxi | 0.304 | 5.38 | ||
| Класс Myxini | 0.237 | 4.23 | ||
| Класс Cephalaspidomorphi | 1.025 | 15.27 | ||
| Класс Chondrichthyes | 1.766 | 27.40 | ||
| Отряд Carcharhiniformes | 1.807 | 26.54 | ||
| Отряд Orectolobiformes | 1.172 | 28.99 | ||
| Отряд Squaliformes | 2.195 | 32.71 | ||
| Отряд Rajiformes | 1.842 | 27.68 | ||
| Отряд Torpediniformes | 0.950 | 23.49 |
| Класс Osteichthyes | 1.799 | 30.14 | ||
| Подкласс Sarcopterygii | 0.570 | 14.10 | ||
| Подкласс Actinopterygii | 1.634 | 24.21 | ||
| Отряд Polypteriformes | 0.653 | 16.15 | ||
| Отряд Lepisosteiformes | 0.984 | 19.49 | ||
| Отряд Acipenseriformes | 1.974 | 30.46 | ||
| Подкласс Teleostei | 1.816 | 30.46 | ||
| Отряд Anguilliformes | 1.208 | 24.98 | ||
| Отряд Cypriniformes | 1.774 | 26.85 | ||
| Отряд Siluriformes | 0.877 | 16.80 | ||
| Отряд Cyprinodontiformes | 1.492 | 32.58 | ||
| Отряд Scorpaeniformes | 1.652 | 29.27 | ||
| Отряд Pleuronectiformes | 1.762 | 29.49 | ||
| Отряд Mugiliformes | 1.411 | 29.95 | ||
| Отряд Gasterosteiformes | 1.739 | 31.60 | ||
| Отряд Perciformes | 1.884 | 33.09 | ||
| Отряд Gadiformes | 2.114 | 33.00 | ||
| Отряд Salmoniformes | 2.331 | 34.72 | ||
| Отряд Gonorhynchiformes | 2.131 | 52.70 | ||
| Отряд Clupeiformes | 3.779 | 56.31 | ||
| Класс Amphibia | 0.719 | 12.79 | ||
| Отряд Caudata | 0.394 | 7.84 | ||
| Отряд Gymnophiona | 0.509 | 12.60 | ||
| Отряд Anura | 0.966 | 16.49 | ||
| Наземные позвоночные | ||||
| Класс Reptilia | 0.861 | 18.22 | ||
| Отряд Rhynchocephalia | 0.510 | 12.62 | ||
| Отряд Testudines | 0.810 | 15.26 | ||
| Отряд Squamata | 0.868 | 18.35 | ||
| Отряд Crocodylia | 0.880 | 21.76 | ||
| Класс Mammalia | 24.29 | 104.00 | ||
| Отряд Monotremata | 8.68 | 77.28 |
| Отряд Marsupialia | 13.46 | 88.33 | ||
| Отряд Sirenia | 9.79 | 54.59 | ||
| Отряд Insectivora | 39.50 | 155.84 | ||
| Сем. Erinaceidae | 12.75 | 59.36 | ||
| Сем. Tenrecidae | 7.79 | 68.15 | ||
| Сем. Soricidae | 47.28 | 180.83 | ||
| Отряд Edentata | 10.11 | 76.44 | ||
| Отряд Lagomorpha | 23.25 | 82.53 | ||
| Отряд Hyracoidea | 14.63 | 82.60 | ||
| Отряд Carnivora | 23.98 | 83.06 | ||
| Отряд Pholidota | 8.58 | 90.59 | ||
| Отряд Tylopoda | 19.08 | 97.69 | ||
| Отряд Rodentia | 24.76 | 100.01 | ||
| Отряд Artiodactyla | 27.33 | 108.47 | ||
| Отряд Pinnipedia | 31.91 | 113.49 | ||
| Отряд Chiroptera | 19.09 | 117.95 | ||
| Отряд Primates | 20.44 | 118.03 | ||
| Сем. Hominidae | 24.41 | 148.17 | ||
| Отряд Cetacea (Сем. Delphinidae) | 34.54 | 118.09 | ||
| Отряд Perissodactyla | 32.28 | 136.87 | ||
| Класс Aves | 32.64 | 111.86 | ||
| Отряд Apterygiformes | 11.88 | 60.28 | ||
| Отряд Caprimulgiformes | 15.91 | 63.12 | ||
| Отряд Gruiformes | 21.03 | 64.75 | ||
| Отряд Ralliformes | 22.32 | 68.99 | ||
| Отряд Sphenisciformes | 22.34 | 70.75 | ||
| Отряд Anseriformes | 24.25 | 71.37 | ||
| Отряд Cuculiformes | 21.07 | 71.68 | ||
| Отряд Columbiformes | 21.93 | 72.73 | ||
| Отряд Falconiformes | 21.90 | 76.90 | ||
| Отряд Casuariiformes | 16.76 | 76.91 | ||
| Отряд Strigiformes | 18.46 | 77.25 |
| Отряд Ciconiiformes | 23.52 | 82.07 | ||
| Отряд Galliformes | 22.75 | 86.36 | ||
| Отряд Piciformes | 27.78 | 92.25 | ||
| Отряд Procellariiformes | 29.07 | 92.78 | ||
| Отряд Trogoniformes | 21.83 | 94.40 | ||
| Отряд Lariformes | 30.43 | 95.62 | ||
| Отряд Coraciiformes | 26.59 | 96.12 | ||
| Отряд Charadriiformes | 29.33 | 101.86 | ||
| Отряд Psittaciformes | 26.57 | 109.19 | ||
| Отряд Rheiformes | 23.65 | 113.99 | ||
| Отряд Struthioniformes | 24.25 | 114.19 | ||
| Отряд Passeriformes | 37.24 | 116.35 | ||
| Сем. Covidae | 32.63 | 107.83 | ||
| Отряд Alciformes | 43.30 | 116.71 | ||
| Отряд Pelecaniformes | 28.29 | 121.33 | ||
| Отряд Apodiformes | 26.52 | 126.91 | ||
| Отряд Coliiformes | 59.74 | 286.08 |
Как видно из этой таблицы коэффициент а и критерий упорядоченности Сr значительно возрастает по мере усложнения строения и функционирования животных. Коэффициент а, представленный в табл.41 характеризует сопоставимый уровень стандартного обмена разных таксонов животных. Его увеличение по мере усложнения строения животных показывает, что прогрессивная эволюция сопровождалась значительным усилением энергетического метаболизма. Так, гребневики имеют стандартный обмен в два раза больший, чем амебы, а моллюски в десять раз больше, чем последние. У позвоночных энергетический метаболизм выше, чем у беспозвоночных, за исключением насекомых, у которых стандартный обмен (коэффициент а) выше, чем у амфибий, рыб и рептилий (табл.41). Среди позвоночных самая большая величина стандартного обмена наблюдается у птиц.
Оказалось также (Зотин, Криволуцкий, 1982; Зотин, 1984; Zotin, 1990), что если сопоставить данные о стандартном обмене отдельных типов и классов животных со временем обнаружения их в палеонтологической летописи, то мы
получаем кривую биоэнергетического прогресса в реальном геологическом масштабе времени. Теперь мы можем уточнить эту кривую, используя данные разных авторов (Друшиц, Обручева, 1971; Геохронологическая таблица, 1974; Savage, Rüssel, 1983; Афанасьев, 1987; Зотин и др., 1990; Расницын, 1990; Кэрролл, 1992, 1993 а,b) о времени возникновения тех или иных таксонов животных в процессе эволюции (табл.42).
Таблица 42
Время возникновения отдельных типов, классов и отрядов животных. Данные о времени возникновения таксонов животных взяты из разных сводок.
| Таксон | Время возникновения таксона | Тcp* | Авторы** | |
| Геологический период | млн. лет | млн. лет | ||
| Тип Sarcomastigophora | Докембрий | |||
| Тип Ctenophora | Докембрий | |||
| Тип Coelenterata | Нижний кембрий | |||
| Класс Scyphozoa | Средний кембрий | |||
| Класс Hydrozoa | Нижний кембрий | |||
| Класс Anthozoa | Нижний ордовик | |||
| Тип Arthropoda | Нижний кембрий | |||
| Класс Myriapoda | Средний кембрий | |||
| Класс Crustacea | Средний кембрий | |||
| Отряд Cirripeda | Силур | |||
| Отряд Mysidacea | Средний карбон | |||
| Отряд Decapoda | Нижний триас | |||
| Отряд Isopoda | Средний триас | |||
| Отряд Tanaidacea | Средняя юра | |||
| Отряд Amphipoda | Средний девон | |||
| Отряд Copepoda | Средний триас | |||
| Отряд Phyllopoda | Средний карбон | |||
| Отряд Ostracoda | Нижний ордовик | |||
| Отряд Anostraca | Средний эоцен | |||
| Класс Arachnida | Силур | |||
| Отряд Amblypygi | Средний карбон |
| Отряд Acariformes | Средний девон | |||
| Отряд Scorpiones | Силур | |||
| Отряд Parasitiformes | Палеоген | |||
| Отряд Aranei | Средний девон | |||
| Отряд Solifugae | Средний карбон | |||
| Отряд Opiliones | Средний карбон | |||
| Класс Insecta | Нижний карбон | 5,6 | ||
| Отряд Collembola | Средний девон | |||
| Отряд Blattodea | Средний карбон | 5,6 | ||
| Отряд Hemiptera | Нижняя юра | |||
| Отряд Dermoptera | Средняя юра | |||
| Отряд Isoptera | Палеоген | |||
| Отряд Plecoptera | Нижняя пермь | 5,6 | ||
| Отряд Orthoptera | Верхний карбон | 5,6 | ||
| Отряд Homoptera | Нижний карбон | |||
| Отряд Odonata | Нижний карбон | 5,6 | ||
| Отряд Coleoptera | Нижняя пермь | 5,6 | ||
| Отряд Lepidoptera | Палеоген | |||
| Отряд Diptera | Верхний триас | 5,6 | ||
| Отряд Hymenoptera | Верхний триас | 5,6 | ||
| Сем. Apidae | Палеоген | |||
| Сем. Formicidae | Палеоген | |||
| Тип Mollusca | Нижний кембрий | |||
| Класс Loricata | Нижний девон | 8,9 | ||
| Класс Bivalvia | Верхний кембрий | 9, 10 | ||
| Отряд Taxodonta | Верхний кембрий | |||
| Отряд Eulamellibranchia | Нижний ордовик | |||
| Отряд Dysodonta | Нижний ордовик | |||
| Класс Gastropoda | Нижний кембрий | 7,11 | ||
| Отряд Gymnosomata | Средний эоцен | |||
| Отряд Basommatophora | Средний карбон | |||
| Отряд Archaegastropoda | Нижний кембрий |
| Отряд Mesogastropoda | Средний ордовик | |||
| Отряд Nudibranchia | Верхний мел | |||
| Отряд Stylommatophora | Верхний мел | |||
| Отряд Neogastropoda | Верхний мел | |||
| Отряд Cephalaspiea | Средний карбон | |||
| Класс Cephalopoda | Нижний кембрий | 12, 13 | ||
| Отряд Nautilida | Нижний кембрий | |||
| Отряд Octopoda | Верхний мел | |||
| Отряд Decapoda | Верхний мел | |||
| Тип Echinodermata | Нижний кембрий | 7, 15 | ||
| Класс Echinoidea | Верхний ордовик | |||
| Класс Asteroidea | Нижний ордовик | |||
| Класс Holothuroidea | Нижний девон | |||
| Класс Ophiuroidea | Нижний ордовик | |||
| Класс Crinoidea | Нижний ордовик | |||
| Тип Tunicata | Верхний силур | |||
| Тип Vertebrata | Нижний кембрий | |||
| Класс Amphioxi | Нижний кембрий | |||
| Класс Myxini | Средний кембрий | |||
| Класс Cephalaspidomorphi | Средний кембрий | |||
| Класс Chondrichthyes | Средний девон | |||
| Отряд Carcharhiniformes | Верхняя юра | |||
| Отряд Orectolobiformes | Верхняя юра | |||
| Отряд Squaliformes | Верхняя юра | |||
| Отряд Rajiformes | Верхняя юра | |||
| Отряд Torpediniformes | Средний эоцен | |||
| Класс Osteichthyes | Средний девон | |||
| Подкласс Sarcopterygii | Средний девон | |||
| Подкласс Actinopterygii | Средний девон | |||
| Отряд Polypteriformes | Средний эоцен | |||
| Отряд Lepisosteiformes | Верхний мел |
| Отряд Acipenseriformes | Нижняя юра | |||
| Подкласс Teleostei | Верхний триас | |||
| Отряд Anguilliformes | Верхний мел | |||
| Отряд Cypriniformes | Верхний мел | |||
| Отряд Siluriformes | Верхний мел | |||
| Отряд Cyprinodontiformes | Средний олигоцен | |||
| Отряд Scorpaeniformes | Палеоцен | |||
| Отряд Pleuronectiformes | Средний эоцен | |||
| Отряд Mugiliformes | Средний эоцен | |||
| Отряд Gasterosteiformes | Средний эоцен | |||
| Отряд Perciformes | Верхний мел | |||
| Отряд Gadiformes | Палеоцен | |||
| Отряд Salmoniformes | Нижний эоцен | |||
| Отряд Gonorhynchiformes | Верхний мел | |||
| Отряд Clupeiformes | Верхний мел | |||
| Класс Amphibia | Верхний девон | |||
| Отряд Caudata | Верхняя юра | |||
| Отряд Anura | Нижняя юра | |||
| Класс Reptilia | Верхний карбон | |||
| Отряд Testudines | Верхний триас | |||
| Отряд Squamata | Верхняя пермь | |||
| Отряд Crocodylia | Мел | |||
| Класс Mammalia | Верхний триас | |||
| Отряд Monotremata | Нижний мел | 16, 19 | ||
| Отряд Marsupialia | Нижний мел | 16, 19 | ||
| Отряд Sirenia | Средний эоцен | |||
| Отряд Insectivora | Верхний мел | 16, 19 | ||
| Сем. Erinaceidae | Нижний эоцен | |||
| Сем. Tenrecidae | Плейстоцен | |||
| Сем. Soricidae | Верхний эоцен | |||
| Отряд Edentata | Верхний палеоцен | |||
| Отряд Lagomorpha | Верхний палеоцен |
| Отряд Hyracoidea | Средний эоцен | 16, 19 | ||
| Отряд Carnivora | Средний эоцен | 16, 19 | ||
| Отряд Pholidota | Верхний палеоцен | |||
| Отряд Tylopoda | Средний эоцен | 16, 19 | ||
| Отряд Rodentia | Нижний эоцен | 16, 19 | ||
| Отряд Artiodactyla | Нижний эоцен | 16, 19 | ||
| Отряд Pinnipedia | Нижний миоцен | 16, 19 | ||
| Отряд Chiroptera | Нижний эоцен | 16, 19 | ||
| Отряд Primates | Палеоцен | 16, 19 | ||
| Сем. Hominidae | Верхний плиоцен | |||
| Отряд Cetacea | Средний эоцен | |||
| Сем. Delphinidae | Нижний миоцен | |||
| Отряд Perissodactyla | Нижний эоцен | 16, 19 | ||
| Класс Aves | Верхняя юра | |||
| Отряд Apterygiformes | Плейстоцен | |||
| Отряд Caprimulgiformes | Средний плиоцен | |||
| Отряд Gruiformes | Средний эоцен | |||
| Отряд Ralliformes | Средний эоцен | |||
| Отряд Sphenisciformes | Средний олигоцен | |||
| Отряд Anseriformes | Верхний мел | |||
| Отряд Cuculiformes | Средний эоцен | |||
| Отряд Columbiformes | Средний олигоцен | |||
| Отряд Falconiformes | Средний эоцен | |||
| Отряд Casuariiformes | Плейстоцен | |||
| Отряд Strigiformes | Средний эоцен | |||
| Отряд Ciconiiformes | Средний эоцен | |||
| Отряд Galliformes | Средний эоцен | |||
| Отряд Piciformes | Верхний эоцен | |||
| Отряд Procellariiformes | Средний эоцен | |||
| Отряд Trogoniformes | Средний эоцен | |||
| Отряд Lariformes | Средний эоцен | |||
| Отряд Coraciiformes | Верхний эоцен |
| Отряд Charadriiformes | Средний эоцен | |||
| Отряд Psittaciformes | Средний олигоцен | |||
| Отряд Rheiformes | Средний плиоцен | |||
| Отряд Struthioniformes | Средний миоцен | |||
| Отряд Passeriformes | Средний эоцен | |||
| Сем. Covidae | Плейстоцен | |||
| Отряд Alciformes | Средний эоцен | |||
| Отряд Pelecaniformes | Верхний мел | |||
| Отряд Apodiformes | Средний эоцен |
Примечания: *) Рассчитано по формуле (129)
**) Авторы: 1 - Несис, 1984; 2 - Соколов, 1962; 3 - Ланге, 1969а; Ланге, 19696; 4 - Чернышева, 1960; 5 - Родендорф, 1962; 6 - Расницын, 1990; 7 - Несис, 1994; 8 - Властов, Матекин, 1968; 9 - Эберзин, 1960; 10 - Филатова, 1968; 11 - Пчелинцев, Коробков, 1960; 12 - Природа, 1984. №1. С. 119; Природа, 1987. №11. С. 116; 13 - Руженцев, 1962; 14 - Луппов, Друщиц, 1958; 15 - Геккер, 1964; 16 - Кэрролл, 1992, 1993а.б; 17 - Обручев, 1964; 18 - Рождественский, Татаринов, 1964; 19 - Громова, 1962.
Следует отметить, что для крупных таксонов время возникновения датируется часто по ныне вымершим группам животных. В то же время коэффициент а и критерий упорядоченности Сr мы расчитывали для современных животных. Поэтому для построения кривых биоэнергетического прогресса и прогрессивной эволюции мы использовали среднее время происхождения тех современных групп животных, по которым рассчитывались коэффициент а и критерий упорядоченности Сr:
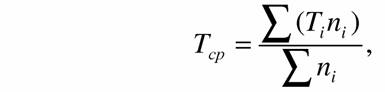
| (129) |
| где Тср - среднее время появления современных животных; Τi - время возникновения таксонов более низкого порядка; ni - число измерений дыхания для соответствующих групп животных. |
На рис.28 показано изменение коэффициента а и критерия упорядоченности Сг в процессе реальной эволюции разных типов животных по данным, приводимым в табл.41 и 42. Как видно из этого рисунка сопоставимый стандартный обмен [коэффициент а из аллометрических соотношений (46) и (47)] непрерывно растет по мере усложнения строения и функционирования животных в процессе эволюции. Здесь интенсивность потребления кислорода пойкилотермными животными приведена к температуре 20°С, а интенсивность дыхания гомойотермных животных приведена к термонейтральной зоне. Кривая, изображенная на рис.28, показывает направление и скорость биоэнергетического прогресса животных.
Если мы хотим построить кривую собственно прогрессивной эволюции, то должны использовать не коэффициент а из аллометрического соотношения (46) или (47), а критерий упорядоченности (74). В состав этого критерия в качестве переменной входит вместо коэффициента а коэффициент а'. В отличие от коэффициента а, коэффициент а зависит, как не раз говорилось, от климатических условий, в которых живут животные, от степени оводненности их тела и других особенностей строения и условий жизни животных. Особо следует подчеркнуть, что при вычислении критерия упорядоченности средние величины коэффициента а надо приводить к температуре 20°С не только для пойкилотермных, но и для гомойотермных. Это делается по единой кривой Крога (68). Другими словами, сравниваемые величины коэффициентов а для млекопитающих и птиц, приводимые в работах Дольника (1968, 1982), и других авторов (Коноплев, Зотин, 1975; Коноплев и др., 1975; Зотин, Криволуцкий, 1982; Зотин, 1984; Зотин и др., 1990; Zotin, 1990) относятся только к биоэнергетическому прогрессу, а не к прогрессивной эволюции.
Как говорилось, биоэнергетический прогресс характеризует увеличение энергетики организмов в процессе эволюции, а прогрессивная эволюция - степень удаленности организмов от начального, близкого к равновесию, состояния среды, в которой зародилась жизнь.
Кривая прогрессивной эволюции для разных типов животных показана на том же рисунке, что и кривая биоэнергетического прогресса (рис.28). Эти кривые, конечно, немного отличаются друг от друга, но это отличие не носит
принципиального характера, и это естественно, так как и та и другая связаны с изменением стандартного обмена в ходе эволюции.
– Конец работы –
Эта тема принадлежит разделу:
А.И.Зотин, А.А.Зотин НАПРАВЛЕНИЕ, СКОРОСТЬ И МЕХАНИЗМЫ ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ
На сайте allrefs.net читайте: "А.И.Зотин, А.А.Зотин НАПРАВЛЕНИЕ, СКОРОСТЬ И МЕХАНИЗМЫ ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Изменение энергетического обмена в ходе эволюции
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.039 сек.
Новости и инфо для студентов