А.И.Зотин, А.А.Зотин НАПРАВЛЕНИЕ, СКОРОСТЬ И МЕХАНИЗМЫ ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ
БИБЛИОГРАФИЯ = Зотин А.И., Зотин А.А. Направление, скорость и механизмы прогрессивной эволюции. – М.: Наука, 1999. – 495 с. - http://www.chronos.msu.ru/relectropublications.html Б-ка электронных публикаций семинара по темпорологии МГУ 31.03.2002.
Зотин А.И., Зотин А.А. Направление, скорость и механизмы прогрессивной эволюции. – М.: Наука, 1999. – 495 с. - http://www.chronos.msu.ru/relectropublications.html Б-ка электронных публикаций семинара по темпорологии МГУ 31.03.2002.
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Российский фонд фундаментальных исследований
Институт биологии развития им.Н.К.Кольцова
А.И.Зотин, А.А.Зотин
НАПРАВЛЕНИЕ, СКОРОСТЬ И
МЕХАНИЗМЫ ПРОГРЕССИВНОЙ
ЭВОЛЮЦИИ
Термодинамические основы
МОСКВА "НАУКА" 1999
Посвящается хирургам
Роберту Мамиконовичу Карапетяну
и
Тимуру Кобаевичу Харатишвили
благодаря искусству которых
данная книга получила свое завершение.
ПРЕДИСЛОВИЕ
Проблемам эволюции животных и растений посвящено огромное количество исследований и казалось бы в этой области биологии все ясно. Однако это не так. Все более или менее ясно в отношении проблем микроэволюции: механизмов видообразования, объяснения целесообразности строения и поведения организмов, приспособленности организмов к условиям существования. Но иначе обстоит дело с пониманием макроэволюционных изменений и, в частности, до сих пор не разработаны в надлежащей мере такие кардинальные проблемы как направленность эволюционного процесса, наличие и механизмы прогрессивной эволюции и др. Конечно, начиная с Ламарка и до наших дней, осуществлено большое число попыток решить проблему эволюционного прогресса и регресса (см. Завадский, 1973; Филипченко, 1977; Назаров, 1984), но при этом большинство ученых (не считая дарвинистов, которые, фактически, отмахнулись от решения указанных проблем) опирались на представление о существовании в живой природе особых сил, отличных от сил и законов остального физического мира. Такой разрыв единой Природы на отдельные компартменты, в которых выполняются разные законы, вызывал и вызывает сопротивление, тем более, что предлагавшиеся особые силы и законы, как правило, носили качественный, словесный характер, что резко снижает их ценность по сравнению с известными физическими законами.
В то же время нет сомнений, что существует специфика жизненных явлений, заметно отличающая их от процессов, протекающих в неживой природе.
Эта специфика состоит в том, что все без исключения живые системы являются организованными, т.е. системами в которых осуществляются процессы регуляции и управления. Теория организованных систем еще не создана, хотя в таких науках, как информатика, кибернетика и синергетика к настоящему времени достигнут значительный прогресс.
Что касается процесса эволюции организмов, как и любых других биологических процессов, то для понимания многих ее проблем необходимо использовать достижения современной термодинамики. Больше того по мнению Викена (Wicken, 1980, 1987) использование термодинамики неравновесных процессов должно стать парадигмой конца двадцатого века, которая позволяет решать проблемы не решенные и не решаемые дарвинизмом. Дело в том, что термодинамика является общей феноменологической теорией любых процессов и явлений, протекающих в природе, сопровождающихся диссипацией энергии. Несомненно, что при протекании любых биологических процессов происходит диссипация энергии, так как они происходят в системах, далеких от равновесия. Поэтому термодинамика может и должна быть использована для решения биологических проблем, в том числе и проблем эволюции.
Однако для действенности провозглашенной Викеном парадигмы необходимо, чтобы, во-первых, была создана термодинамика нелинейных неравновесных процессов и, во-вторых, термодинамика должна быть объединена с науками, занимающимися проблемами организации и самоорганизации, т.е. создана термодинамика организованных систем. То и другое еще далеко от завершения. И все же некоторые достижения в области термодинамики неравновесных процессов и организованных систем имеются и было бы неразумным не использовать их для понимания общих проблем биологической эволюции. В частности, возможно введение на базе термодинамики неравновесных
процессов четкого определения прогрессивной эволюции и получение количественных критериев этого процесса.
В предыдущих двух книгах были рассмотрены термодинамические основы физиологических процессов (Зотин, 1988) и онтогенеза организмов (Зотин, Зотина, 1993). Настоящая монография посвящена проблемам биологической эволюции. В основу рассмотрения проблем макро и мегаэволюции и особенно прогрессивной эволюции в данной книге положены два термодинамических принципа: наименьшей диссипации энергии и наискорейшего спуска. Первый из них определяет общее направление эволюции любых термодинамических систем (в том числе и живых организмов), второй - возникновение в процессе эволюции высокоупорядоченных и организованных живых систем.
Охватить все вопросы эволюционного учения, опираясь на термодинамику, нереально, хотя термодинамический подход к проблемам эволюции пользуется известной популярностью среди специалистов (см. Wicken, 1987; Brooks, Wiley, 1988; Гладышев, 1988, 1996; Leuschner, 1989; Swensen, 1989, 1992, 1997; Хазен, 1993 a, 1996). Мы коснемся указанных исследований очень кратко и лишь в той части, где они решают проблемы макро- и мегаэволюции, направленности эволюционных процессов, прогрессивной и регрессивной эволюции. Следует отметить при этом, что термодинамические соотношения будут приведены и использованы в минимальной степени - их более полное изложение приведено в предыдущих книгах (Зотин, 1988; Zotin, 1990) и статьях (Зотин, Зотин, 1995; Zotin, Zotin, 1996). Несмотря на это, данная монография, как и предшествующие пронизана термодинамической методологией, так как по нашему убеждению, как и по убеждению многих других авторов, теоретической основой любого макроскопического биологического процесса являются термодинамические закономерности.
В монографии использованы многочисленные экспериментальные данные, полученные в Лаборатории биофизики развития Института биологии развития РАН. С сотрудниками этой лаборатории осуществлялась не только совместная экспериментальная работа, но и проводились многочисленные дискуссии по обсуждаемым в книге проблемам. Всем участникам этих работ и дискуссий и особенно Т.А.Алексеевой, И.Г.Владимировой, В.В.Гаврилову, В.А.Грудницкому, С.Ю.Клейменову, В.А.Коноплеву, И.С.Никольской, Н.Д.Озернюку, Е.В.Преснову, Л.И.Радзинской, В.С.Фаустову, а также профессорам И.Лампрехту, Б.Шааршмидту и Д.Лёйшнеру (Германия) мы приносим свою искреннюю благодарность. Мы благодарим также директора Института биологии развития РАН, академика Н.Г.Хрущева, профессоров Б.Р.Стриганову и С.Г.Васецкого за благожелательную критику данной работы. Книгу удалось опубликовать только благодаря помощи члена-корреспондента РАН Г.И.Абелева и финансовой поддержке Российского фонда фундаментальных исследований.
Часть 1. ТЕРМОДИНАМИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИИ
Зотина, 1993; Зотин, Зотин, 1995; Zotin, Zotin, 1996). Соответственно в данной книге принимается, что многие проблемы макро- и мегаэволюции можно…Глава 1. Необходимые сведения из термодинамики неравновесных процессов
Подробное изложение основных методов и соотношений термодинамики неравновесных процессов можно найти в многочисленных руководствах и учебниках (Денбиг, 1954; де Гроот, 1956; Пригожин, 1960; де Гроот, Мазур, 1964; Хаазе, 1967; Залевски, 1973; Дьярмати, 1974; Бахарева, 1976; Преснов, 1976а; Гуров, 1978; Журавлев, 1979; Leuschner, 1979; Быстрай, 1987) и специальных обзорах и монографиях по термодинамике биологических процессов (Антонов, 1968; Зотин, 1974, 1976; 1988; Николаев, 1976; Рубин, 1984; Кеплен, Эссиг, 1986; Brooks, Wiley, 1988; Leuschner, 1989; Зотин, Зотина, 1993). Ниже мы приведем лишь краткие выводы из этих работ, опираясь, естественно, в основном на наши публикации (Зотин, 1988, Zotin, 1990, Зотин, Зотина, 1993; Зотин, Зотин, 1995, Zotin, Zotin, 1996).
В основе термодинамики неравновесных процессов, кроме соотношений классической термодинамики, лежат следующие положения:
1. Линейные законы, согласно которым вблизи от равновеси
выполняются феноменологические уравнения

|
| 2. Соотношение взаимности Онзагера |
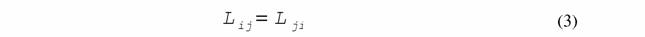
|
| или в нелинейной области |
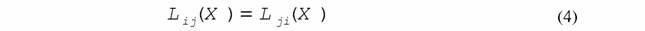
|
| 3. Разбиение скорости изменения энтропии на два члена |
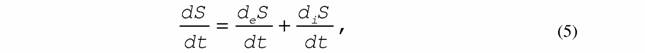
|
где deS / dt - скорость поступления или выхода энтропии из открытой системы, которая носит название потока энтропии, а diS / dt - продукция энтропии внутри системы.
| 4. Второе начало термодинамики в виде |

|
| показывающее, что любой необратимый процесс, протекающий внутри системы, сопровождается возникновением энтропии, т.е. диссипацией энергии. 5. Уравнение Гиббса |
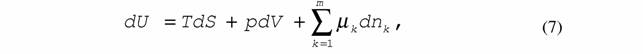
|
| где U - внутренняя энергия системы; Т - абсолютная температура; S - энтропия; p - давление; V - объем; μk - химический потенциал; n - число молей вещества k. Уравнение (7) особенно широко используют в термодинамике линейных необратимых процессов. 6. Конститутивные уравнения |
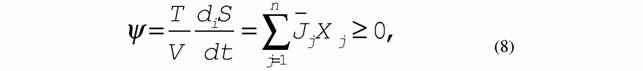
|
| где ψ- удельная диссипативная функция системы. В нелинейной области оно имеет вид |
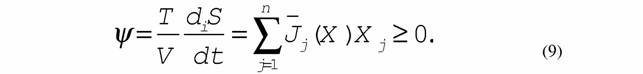
|
11 7. Сопряженные процессы:
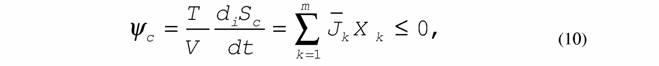
|
| которые осуществляются только при обязательном выполнении неравенства (8) или (9). 8. Принцип минимальной скорости продукции энтропии в стационарном состоянии (теорема Пригожина): |
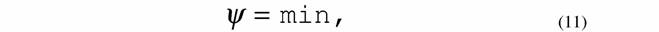
|
| из которой следует критерий эволюции: |

|
| Критерий эволюции (12) показывает, что в процессе релаксации системы к равновесному или стационарному состоянию удельная диссипативная функция непрерывно уменьшается пока не достигнет минимальной величины (11). 9. Критерий эволюции для нелинейных систем, близких к линейным, вида (Зотин, Зотина, 1977; Зотина, Зотин, 1980; Зотин, 1988; Leuschner, 1989; Завальнюк, 1991): |
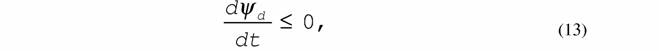
|
| где Ψd - функция внешней диссипации (псидфункция), которая введена исходя из следующих соображений. Известно (Пригожин, 1960), что для химических систем справедлива формула |
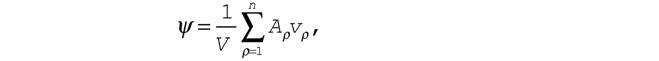
|

|
| где G - свободная энергия Гиббса; H - энтальпия. Подставляя это выражение в предыдущее, получаем |
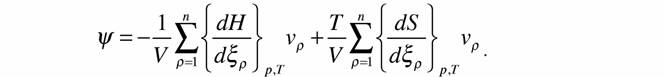
|
| Теперь, вводя обозначения: |
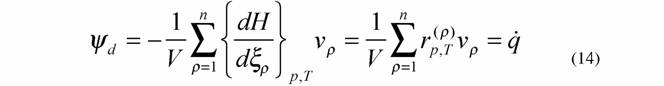
|
| и |
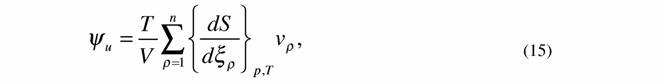
|
| получаем разбиение |
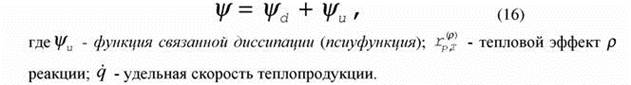
|
Разбиение (16) вытекает также из рассмотрения физического смысла диссипации энергии в открытой системе (Зотин, 1974) и из различных термодинамических соображений (Landauer, 1973; Tykodi, 1974; Wangsness, 1975; Lurie, Wagensberg, 1979a,b; Leuschner, 1981, 1989; Завальнюк, 1991). Приведенное
выше доказательство разбиения (16) справедливо не только для чисто химических, но и для любых других термодинамических систем (Зотина, Зотин, 1980).
Физический смысл функции внешней диссипации ясен (14), поэтому, критерий эволюции (13) может быть записан в виде

|
| Таким образом, критерий эволюции, как для систем близких к равновесию (12), так и для слабо нелинейных систем (13), показывает, что в процессе изменения этих систем происходит уменьшение интенсивности диссипации энергии, которая в стационарном состоянии достигает минимальной величины. Для сильно нелинейных систем доказать неравенства типа (12) и (13) пока не удается, но можно их ввести, опираясь на следующий постулат. 10. Принцип наименьшей диссипации энергии, согласно которому (Зотин, 1988, Зотин, Зотина, 1993): в устойчивом состоянии любой термодинамической системы скорость диссипации энергии в ней минимальна т.е. |
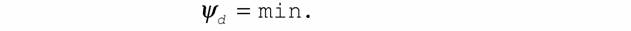
|
| Если речь идет о сильно нелинейных системах, то в этом случае псидфункция в стационарном состоянии не обязательно константна и критерии эволюции (13), (17) можно записать как |
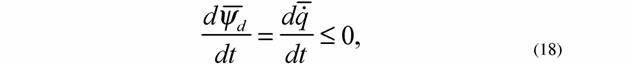
|
| а принцип наименьшей диссипации энергии в виде |
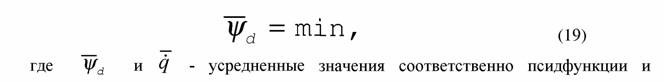
|
| интенсивности теплопродукции системы. |
Н.Н.Моисеев (1987) предложил несколько иную формулировку принципа наименьшей диссипации энергии, которая звучит следующим образом: "если допустимо не единственное состояние системы (процесса), а целая совокупность состояний, согласных с законами сохранения и связями, наложенными на систему (процесс), то реализуется то ее состояние, которому отвечает минимальное рассеяннее энергии, или что то же самое, минимальный рост энтропии" (стр.27). По сути дела эта формулировка мало отличается от приведенной выше. Другими словами, Моисеев (1987) и мы (Zotin, Zotina, 1971; Зотин, Зотина, 1976, 1993; Зотин, 1974, 1988) независимо друг от друга предложили новый термодинамический принцип, который включает в себя второе начало и охватывает практически все явления природы. Следует, однако, отметить, что, переходя к биологическим проблемам, Н.Н.Моисеев несколько занизил значение предложенного принципа, сведя его фактически к принципу оптимальности.
Принцип наименьшей диссипации энергии можно графически изобразить в виде схемы, показанной на рис. 1(1). Положение шарика на дне чаши с круглым дном соответствует наиболее устойчивому, стационарному состоянию системы. Если систему вывести из этого состояния, то она стремится вернуться в прежнее устойчивое состояние с минимальной интенсивностью диссипации энергии. Подобного рода изменения можно назвать нестационарным переходным процессом (Зотин, 1988).
11. Интегральный принцип наименьшей диссипации энергии, согласно которому (Зотин, Алексеева, 1984; Зотин, Зотина, 1987, 1993):
устойчивым является такой неравновесный процесс, при котором суммарная удельная диссипация энергии в системе за изучаемый отрезок времени минимальна
Фактически это утверждение является следствием принципа наименьшей диссипации энергии (19), и его можно записать в следующем виде:
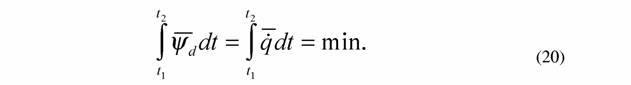
|
Неравновесный процесс типа (20) осуществляется через ряд
стационарных состояний, т.е. путем стационарного переходного процесса. Такой процесс можно изобразить в виде схемы, показанной на рис.2(1).
Как отметил Пригожин (1959, 1960), если в системе или в системе и среде имеется двойной масштаб времени протекания необратимых процессов, то переход системы к состоянию равновесия само собой разбивается на два этапа: появление стационарного состояния и эволюция системы через ряд стационарных состояний к равновесию. Наиболее устойчивая траектория (или креод по терминологии Уоддингтона, 1970) проходит по дну корыта [рис.2(1)].
Вероятностная интерпретация критериев эволюции.
вероятностной интерпретации понятия энтропии и диссипативных функций. Ниже мы остановимся только на последнем, так как для характеристики… Процесс приближения термодинамической системы к равновесному или стационарному… Как известно (Гленсдорф, Пригожин, 1973; Залевски, 1973; Гуров, 1978), возникновение флуктуации в изолированной…Кинетика приближения термодинамической системы к конечному состоянию.
системы в начальный момент времени ί=0. Подставляя (30) и (31) в (24) и учитывая разбиение (16), имеем … Это уравнение хорошо согласуется с зависимостью,… времени t=0. Подставляя это решение в (29), имеем Теперь мы можем…Глава 2. Необходимые сведения из теории биологической эволюции
К середине прошлого века оформились в окончательном виде два подхода в изучении эволюции природных систем, сыгравшие выдающуюся роль в науке: теория эволюции физических систем, опирающаяся на второе начало термодинамики (Клаузиус, 1934), и теория эволюции живых систем, опирающаяся на теорию естественного отбора (Уоллес, 1898; Дарвин, 1939). Хотя та и другая теории рассматривали вопрос об эволюции, они резко отличались друг от друга (Гленсдорф, Пригожин, 1973). В первом случае речь шла об эволюции, как о процессе приближения физической системы к состоянию равновесия и связанному с этим упрощению и разрушению системы, во втором - об увеличении сложности биологических систем, которое неизбежно должно было сопровождаться
удалением от состояния равновесия. Отличались и методология исследований. Термодинамика является дедуктивной наукой и ее основной задачей является изучение общих причин и направления изменений любых процессов, протекающих в Природе. В противоположность этому теория естественного отбора возникла на базе индуктивных представлений и стремилась объяснить, главным образом, механизмы биологической эволюции.
Хотя эволюция жизни на Земле - единый процесс, начинающийся с наследственных изменений отдельных организмов и кончающийся преобразованием биоценозов, он складывается из отдельных крупных этапов, каждый из которых имеет свои особенности и свои специфические механизмы. Большинство исследователей выделяют два этапа эволюции: микроэволюцию (возникновение новых видов) и макроэволюцию (последующее появление более крупных таксонов вплоть до типов). Последовательные дарвинисты считают, что способы эволюционных изменений в обоих случаях мало отличаются друг от друга. Однако существует и другая точка зрения, согласно которой макроэволюция имеет свои особенности и механизмы, отличные от механизмов микроэволюции. Более того, кроме понятия макроэволюции выделяют еще и понятие мегаэволюции (изменение биоценозов и всех организмов биосферы в целом), так как она, как будет видно из последующего, также обладает своими специфическими механизмами и особенностями.
Микроэволюция
условиям существования, возникновение целесообразных морфологических структур и поведения. Основой этих пусковых механизмов является естественный… Теория естественного отбора, объясняющая каким образом возникают новые виды,…Аксиоматика
Наличие естественного отбора среди организмов, населяющих Землю, вытекает из следующих очевидных, т.е. взятых из повседневного опыта, аксиом или… Постулат первый:все живые организмы способны размножаться в геометрической… Этот постулат очевиден для одноклеточных организмов, так как простое деление клеток происходит в геометрической…Синтетическая теория эволюции
Последующее развитие науки привело к установлению дискретности наследственных факторов - генов, их малой изменяемости, насыщенности природных… Тимофеев-Ресовский и др., 1969; Шмальгаузен, 1969; Медников, 1973; Левонтин,… 1) географическая изоляция части популяции, что приводит к несколько различным условиям действия естественного отбора…Макроэволюция
Биологический и морфо-физиологический прогресс
возникали все более совершенные и сложно устроенные животные и растения. Направленность эволюционного процесса в этом отношении, т.е. наличие… Прежде всего это касается работ А.Н.Северцова (1925, 1934, 1939), который… Биологический прогресс той или иной группы организмов характеризуется увеличением численности, расширением ареала…Прогрессивная эволюция
случае это касается млекопитающих, шла не так уж быстро, хотя направленность ее очевидна (рис.5). Большинство российских авторов считают, что прогрессивная эволюция равнозначна… Обычно считается, что направленность эволюции отдельных групп животных и растений связана с наличием целого ряда…Биоэнергетический прогресс
на которой можно построить количественные критерии эволюционного прогресса, пригодные для сравнительно-эволюционных исследований. В частности, Ивлев… потребления кислорода; M - масса организма (в г);… Исходя из этого, Ивлев (1959) в качестве основного показателя энергии жизнедеятельности животных предложил…Совершенствование регуляции и управления
повышение энергетического уровня жизненных процессов; 2) повышение эффективности размножения; 3) улучшение точности восприятия внешних сигналов и… Иорданский (1990), исходя из работ Дарвина (1939), Северцова (1925, 1939),… Для характеристики прогрессивной эволюции предложены и некоторые другие понятия, заимствованные, в частности, из…Мегаэволюция
Мегаэволюцией Симпсон (1948), как и некоторые другие палеонтологи, называет образование таксонов высших категорий таких как отряды, классы и типы.… Понятие биосфера формировалось постепенно, начиная с работ Ламарка… Как следует из работ Вассовича и Иванова (1983), Шиловой (1983) и Колчинского (1990) термином биосфера [введенным…Механизмы мегаэволюции.
этом, что изменить сложившийся биоценоз не так-то просто: в результате действия стабилизирующего отбора, теорию которого подробно разработал… Типичные колебательные изменения биогеоценозов связаны с изменением… Иначе обстоит дело с макроэволюционными процессами. Как правило, макроэволюционные изменения связаны с постоянно…Регрессивная эволюция.
Согласно принципу наименьшей диссипации энергии в процессе эволюции любой термодинамической системы происходит ее приближение к равновесному… Советские историки дарвинизма (Завадский, 1958; Завадский, Ермоленко, 1966;… Кратко остановимся на теориях регрессивной эволюции, опираясь, главным образом, на историю этих идей, приводимую в…Глава 3. Термодинамика и биология
Вопрос об использовании термодинамики в биологии всегда вызывал ожесточенные споры (см. Лазарев, 1945; Touloukian, 1956; Wilkie, 1960; Пасынский, 1963; Тринчер, 1965, 1966; Зотин, 1971; Аршавский, 1982 и др.), однако нам, как и многим другим (Блюменфельд, 1977; Зотин, 1974, 1988; Николаев, 1976; Волькенштейн, 1978; Николис, Пригожин, 1979; Wicken, 1979, 1986, 1987; Wiley, Brooks, 1982, 1983; Brooks, Wiley, 1984;, 1988; Рубин, 1984; Leuschner, 1989, Гладышев, 1996), кажется, что его следует решить положительно. Викен (Wicken, 1979, 1986, 1987) даже считает, что использование термодинамики неравновесных процессов для понимания проблем эволюции является новым переломным моментом, новой парадигмой в развитии эволюционного учения после Дарвина. Мы согласны с ним, и в данной книге принимается, что многие проблемы макро- и мегаэволюции, решение которых недоступно дарвинизму, следует рассматривать, опираясь на соотношения и методологию термодинамики неравновесных процессов.
Как известно, термодинамика является наукой о явлениях и процессах, протекающих в материальных системах и сопровождающихся образованием, переносом и превращением тепла. Процессы, происходящие в организмах и с организмами, сопровождаются интенсивным образование тепла в результате диссипации энергии, и к ним, поэтому, применимы термодинамические
закономерности. Это было осознано довольно давно - фактически, с момента возникновения термодинамики - и попытки ее использования в биологии известны с давних времен (см. Лазарев, 1945; Гельфер, 1969, 1973; Брода, 1978).
Дело, однако, осложнялось тем, что до 50 годов нашего века термодинамика была разработана только для изолированных систем, находящихся в состоянии равновесия или квазиравновесия. Живые системы не являются изолированными системами и не находятся в состоянии равновесия. Более того, они очень далеки от такого состояния, т.е. являются сильно неравновесными, открытыми системами (Зотин, 1971). Поэтому классическая термодинамика применима к ним только в ограниченных пределах. В результате, хотя термодинамические расчеты получили достаточно заметное применение в физической биохимии (см., например, Eyring et al., 1960; Пасынский, 1963; Маршалл, 1981, Джоунс, 1982; Кеплен, Эссиг, 1986), неравновесные процессы, которые представляют наибольший интерес для биологии, не могли быть изучены на базе классической термодинамики.
Только с разработкой в трудах де Донде, Онзагера и Пригожина (de Bonder, 1931; Onsager, 1931a,b; Prigogine, 1947, Пригожин, 1960) термодинамики линейных необратимых процессов стало возможным использование ее соотношений для описания некоторых биологических явлений. Здесь были достигнуты заметные успехи не только в отношении частных физиологических процессов (Katchalsky, Curran, 1965; Антонов, 1968; Glaser, 1876; Рубин, 1984), но и более общих, таких как развитие организмов (Зотин, 1974; Рубин, 1984; Brooks, Wiley, 1988; Leuschner, 1989; Зотин, Зотина, 1993), динамика популяций и изменения экосистем (Приц, 1974; Николис, Пригожин, 1979; Рубин, 1984; Brooks, Wiley, 1988; Ulanowicz, 1986), биологическая эволюция (Prigogine et al., 1972; Коноплев, Зотин, 1975; Зотин, Коноплев, 1976; Николис, Пригожин, 1979; Зотин, Криволуцкий, 1982; Зотин, 1984; Brooks, Wiley, 1988; Wicken, 1987; Leuschner, 1989; Swensen, 1989, 1992, 1997; Zotin, 1990; Хазен, 1993; Зотин, Зотин, 1995; Zotin, Zotin, 1996 и др.).
Мы остановимся подробнее только на проблемах биологической эволюции, да и то позднее, а сейчас нам хотелось бы предостеречь от слишком
оптимистического отношения к возможности использования термодинамики для толкования биологических явлений. Соотношения термодинамики линейных необратимых процессов, которая, как и классическая термодинамика, к настоящему времени является завершенной наукой, недостаточны для рассмотрения указанных биологических явлений. Для этого необходимо построение термодинамики организованных систем, включающей термодинамику нелинейных процессов, которые пока еще не созданы в достаточно полном виде и носят во многом субъективный характер. Это не означает, что положения этих разделов термодинамики нельзя использовать в биологии, но не исключено, что многие из них будут значительно изменены в будущем.
Теперь кратко о принципиальных вопросах, связанных с основными понятиями термодинамики и их применимости в биологии.
Энтропия и негэнтропия
Энтропия.
Гельфер, 1969, 1973). Сложность проблемы, если речь идет об использовании этого понятия в биологии, состоит в том, что, во-первых, оно определено… Мы не будем приводить выкладки, с помощью которых получено это выражение, их… Это увеличение энтропии имеет простой физический смысл. Согласно второму началу термодинамики любой неравновесный…Негэнтропия.
негэнтропия —> информация —> негэнтропия (48) Связь между информацией и энтропией в этом случае определяется тем, что для… Попытка связать термодинамику с теорией информации, естественно, вызвала большой интерес у биологов и физиков (см.…Организованные системы
способности к организации. С этой точки зрения, понятие негэнтропия характеризует организованность живой системы. Согласно Поплавскому (1981) первый член в утверждении (48) можно… где ΔST - изменение энтропии термостата, в который помещена изучаемая система;…Живые системы.
составу, но не организованной системе [рис. 1(2)]. Это связано с тем, что организованность клеток, зародышей и целых организмов возникает в результате… Из указанных негэнтропийных эффектов реально измеримым в настоящее время является только интегральный негэнтропийный…Термодинамические подходы к проблемам биологической эволюции
Больцман (1970) пошел еще дальше, высказав идею, что борьба за существование является борьбой за энтропию, которой можно располагать при переходе… Были, конечно, и раньше, и в настоящее время попытки более основательного… В последние годы, как уже отмечалось, для понимания закономерностей и направленности биологической эволюции многие…Термодинамика и направленность эволюции
1979-1987; Wiley, Brooks, 1983; Brooks, 1984; Brooks, Wiley, 1984, 1988; Swensen, 1989, 1992, 1997). Первая проблема, которая встает при попытке использования термодинамики при… К сожалению, многие из них опираются при этом на методологию классической термодинамики, хотя и используют иногда…Общие соотношения биологической термодинамики
выделить четыре типа изменений, определяющих функцию внешней диссипации отдельного организма (Lamprecht, Zotin, 1978; Brooks, Wiley, 1988; Зотин,… где усредненные функции внешней диссипации:-… в то время как для популяций может выполнятся, как такое же, так и обратное неравенство: (62) …Часть 2. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: УСИЛЕНИЕ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Несмотря на то, что Ламарк был ярко выраженным материалистом, его представления об эволюционном прогрессе имели виталистический оттенок (как и у… эволюционного учения после Дарвина, в понимании причин и механизмов…Глава 1. Критерии биоэнергетического прогресса и прогрессивной эволюции
Как отмечалось, формула (14) показывает, что функция внешней диссипации организмов равна удельной теплопродукции живой системы, т.е. реальной диссипации энергии, происходящей в организме. С другой стороны, по энергетическому метаболизму можно судить о потенциальной возможности живой системы диссипировать энергию, так как дыхание и гликолиз соответствуют подводимой к системе мощности. Следовательно, для живых систем должно быть справедливо равенство (Зотин, 1988)
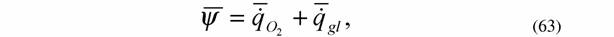
|
где 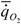 - интенсивность потребления кислорода (скорость потребления кислорода на единицу массы организма); - интенсивность потребления кислорода (скорость потребления кислорода на единицу массы организма);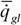 - интенсивность гликолиза.
В аэробных условиях у большинства животных величина - интенсивность гликолиза.
В аэробных условиях у большинства животных величина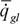 пренебрежимо мала и выражение (63) можно упростить до пренебрежимо мала и выражение (63) можно упростить до
|
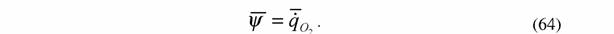
|
При 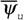 — 0 из (16) следует, что
— 0 из (16) следует, что 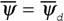 , а так как
, а так как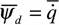 (14), то
(14), то
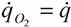 (65)
(65)
где 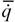 - интенсивность теплопродукции организмов.
- интенсивность теплопродукции организмов.
Энергетический обмен выражают в разных единицах, но в последние годы все чаще в Вт/кг или мВт/г. Этих единиц мы будем придерживаться в книге, пересчитывая данные других авторов, когда это необходимо, используя оксикалорийный коэффициент равным 4,821 кал/мл О2 (Briedis, Seagrave, 1984) и коэффициент 1,163 мВт-час/кал, если данные выражены в кал/час.
Стандартный обмен
В дальнейшем мы будем пользоваться главным образом понятием стандартного обмена, так как этот вид энергетического обмена, с одной стороны, хорошо… Очевидна условность и неточность определения стандартного обмена, как… мелкие животные (простейшие, ракообразные, насекомые) не могут достаточно долгое время (по сравнению с временем…Значение массы тела
Из формулы (46) следует, что для вычисления коэффициента а (при известных значениях и М) необходимо знать величину коэффициента k. По поводу этого коэффициента существует большая литература. Еще Рубнер… Определение коэффициента k требует большого числа измерений дыхания у организмов разной массы. Следует учитывать при…Влияние температуры
Пойкилотермные животные. Проводить измерения дыхания при 20°С не всегда возможно, так как многие из пойкилотермных животных живут при более низких… (67) Естественно, что при получении кривой Крога (зависимости дыхания от температуры), потребление кислорода следует…Акклимация и акклиматизация
С последним, однако, нельзя согласится без специальной экспериментальной проверки. Во всяком случае, полученные к настоящему времени данные на… Второй проблемой, которую необходимо учитывать при определении критерия… теплых поясов Земли (Schölander et al., 1953; Ивлева, 1972, 1981;Критерий упорядоченности
С термодинамической точки зрения состояние равновесия соответствует полному беспорядку - наиболее вероятному состоянию системы. Если система… В классической термодинамике мерой беспорядка является энтропия и ее… сопровождается увеличением неупорядоченности системы.Максимальный обмен
Как и в случае стандартного обмена, максимальный обмен животных сильно зависит от массы тела животного и от температуры среды. Многими авторами… Броди (Brody, 1945), по- видимому, одним из первых предположил, что… На рис.12 приведены данные об соотношении , максимального () и стандартного () обмена у 160 видов насекомых, рыб,…Константа Рубнера
В процессе развития, роста и старения организмов происходит снижение стандартного обмена, прерываемое на некоторых стадиях эмбрионального и раннего… (83) где Zmax - максимальная продолжительность жизни животного (годы); средняя интенсивность потребления кислорода (мВт/г);…Глава 2. Стандартный обмен и критерий упорядоченности у беспозвоночных животных
Как отмечалось, В.Р.Дольник (1968) показал, что сопоставимый стандартный обмен [коэффициент а из аллометрических соотношений (46), (47)] заметно возрастает по мере увеличения сложности организации животных от простейших до млекопитающих и птиц (табл.1). Еще интересней оказалась попытка сопоставления величины стандартного обмена отдельных групп животных с временем возникновения этих групп в реальном палеонтологическом времени (рис.6). Данные, приводимые на рис.6 показывают, что в процессе реальной эволюции шел непрерывный биоэнергетический прогресс.
Для характеристики направления прогрессивной эволюции важнее, однако, понятие критерия упорядоченности, так как, как говорилось, только этот
критерии показывает насколько изучаемая группа животных далека от состояния равновесия, от первичного бульона, в котором зародилась жизнь. Критерий упорядоченности показывает направление прогрессивной эволюции организмов и поэтому он может быть положен в основу макроэволюции и макросистематики животных и растений.
Очевидно, что для количественного описания биоэнергетического прогресса и прогрессивной эволюции важно как можно точнее определить коэффициент а в разных группах животных. В настоящее время в отношении некоторых групп животных это сделать можно, так как в литературе имеется достаточно большое число измерений стандартного обмена представителей разных типов и классов животных. При вычислении критерия упорядоченности, как отмечалось, необходимо также учитывать содержание воды в теле животных [т.е. использовать формулы (76)-(81), а не (75)] и температурную, метаболитическую компенсацию. Из-за сравнительной малочисленности данных это не всегда удается сделать, но стремиться к этому необходимо для более правильного определения критерия упорядоченности.
Специально для этой книги мы еще раз проверили значения величин аллометрических коэффициентов и рассчитали критерий упорядоченности для типов и классов животных, большинство из которых входят в табл.1, а также собрали данные по группам, не рассматривавшихся раньше.
Как говорилось, имеются различные трудности при сопоставлении данных, полученных разными авторами. Это вносит некоторую неуверенность в отношении правильности определения коэффициента α для различных групп животных. Эту неуверенность можно уменьшить только увеличением числа используемых данных, так что по мере использования новых публикаций о исследованиях стандартного обмена и критического анализа старых работ неточность определения величины α и, соответственно, критерия упорядоченности Cr, y разных видов животных должна уменьшаться.
В соответствии с мнением некоторых зоологов и, исходя из удобств изложения, мы разбили всех беспозвоночных животных на три группы: простейших, низших беспозвоночных и высших беспозвоночных. Каждая из этих групп включает в себя несколько типов животных, но все они значительно
отличаются от позвоночных животных не только морфологическим строением, но и количеством имеющихся в литературе данных о стандартном обмене. Поэтому мы выделили тип позвоночных животных в отдельную главу. Изложение мы начнем с материалов о стандартном обмене животных группы простейших (иногда их возводят в ранг подцарства).
Простейшие
Содержание воды в простейших равно в среднем 86.5% (Владимирова, Зотин, 1983). Эту цифру мы приняли для водных организмов в качестве эталонной… Зависимость скорости потребления кислорода от массы тела животных описывается… ориентироваться на данные, приводимые в работах ВладимировойApicomplexaSarcomastigophoraCiliophora
EumycetozoeaZoomastigophoreaLobosea Phytomastigophorea. а в типе Ciliophora: PolymenophoreaOligohymenophoreaKinetofragminophoreaНизшие беспозвоночные
Тип Porifera(Губки) В отношении представителей типа Porifera в литературе имеются лишь единичные… Яковлева, 1963; Виленкина, Виленкин, 1969; Cotter, 1978).ScyphozoaHydrozoaAnthozoa
Для всего типа кишечнополостных коэффициент а (при k=0.75) и критерий упорядоченности Сr, вычисленный с учетом степени оводненности представителей разных классов и температурной компенсации, равны:
Тип Coelenterataa =0.297±0.036; Cr =5.07±0.50 (89)
Черви - Plathelminthes (Плоские черви)
которую мы и использовали для приведения данных авторов к температуре 20° С. … На рис. 14(3) показана кривая зависимости скорости потребления кислорода от массы тела плоских червей по данным,…Высшие беспозвоночные
О дыхании представителей некоторых типов высших беспозвоночных в литературе имеется не много данных. Относительно же членистоногих и моллюсков… Тип Arthropoda(Членистоногие) Тип членистоногих включает в себя несколько классов животных, из которых мы рассмотрим меростомовых, многоножек,…Acariformes —>Amblypygi —> Scorpiones —> Parasitiformes —> —> Aranei —> Solifugae —> Opiliones
Коэффициент а (мВт/г) и критерий упорядоченности Сr в разных таксонах класса Arachnida (Паукообразные). N - число видов, n - число измерений (Зотин… Таксон N n a Cr Отряд… Подотряд Trombidiformes 0,573±0,159 9,84±2,73 Сем.…Низшие насекомые - Apterygota
k=0.75, можно вычислить по формуле (76) значение коэффициента а для Ctenolepisma sp. (a = 0.74) и для L. saccharina (a = 0.31). Отсюда можно принять… Отряд Thysanuraa=0.53; Cr=8.72 (96) Для отряда Collembola (Ногохвостки) получено значительно больше данных (табл.15). Мы рассчитали по данным, приводимым…Высшие насекомые - Pterygota
Как было показано в табл.1 и на рис.6, насекомые достаточно резко выделяются среди других пойкилотермных животных по уровню стандартного обмена. В…Насекомые с неполным превращением
Данных о дыхании тараканов сравнительно немного. В работе Донцовой и др.(1985) приведены собственные и литературные материалы о массе тела и… обмене тараканов представлены в табл.16. Расчет указанных авторов (Алексеева и…Насекомые с полным превращением
Для этого отряда имеются данные о массе тела и дыхании (117 видов насекомых, 133 измерений) из 19 семейств (Алексеева, Зотин, 1995). Авторы приводят… Более точные расчеты дают величину k=0.730±0.050, достоверно не отличающуюся от константы Хемингсена (k=0.75). Мы,…Суперорганизмы.
которые обладают свойствами гомойотермных организмов, т.е. способны к поддержанию постоянной и достаточно высокой температуры внутри этого… Механизм терморегуляции пчел связан с усилением или ослаблением активных… Иначе происходит терморегуляция муравейника. Она осуществляться не за счет повышения или понижения энергетического…Макросистематика насекомых
В настоящее время отсутствуют общепризнанные критерии порядка расположения отрядов в классе насекомых. Это видно хотя бы по тому, как менялось… комбинированный путь построения макросистемы: когда очевиден признак выделения… Таблица25.Taxodonta —> Eulamellibranchia —> Dysodonta
Таблица 27. Масса тела М(г), скорость потребления кислорода (мВт) при 20°С, а (мВт/г) и… (Двустворчатые моллюски). N - число изученных видов, n - число измерений (Зотин, Зотин, 1998).Gymnosomata —> Sacoglossa —> Nudibranchia—> —> Cephalaspidae
Класс Cephalopoda(Головоногие) Головоногие моллюски выделяются среди всех моллюсков и беспозвоночных… Этому соответствует и высокий уровень сопоставимого стандартного обмена - величина коэффициента а из соотношения (46).…Bivalvia—> Gastropoda—> Loricata—> Cephalopoda
Тип Echinodermata (Иглокожие) Работ, в которых изучалась скорость потребления кислорода взрослыми формами… Пересчет данных Рябушко о зависимости дыхания от температуры в типе Echinodermata показал, что в относительном…Echinoidea —>Asteroidea —> Holothuroidea —> —> Ophiuroidea—> Crinoidea
Коэффициент а и критерий упорядоченности Сr для разных классов типа Echinodermata.По данным, представленным в табл.30. N - число изученных видов. … Класс а Сr N Echinoidea… Holothuroidea (Голотурии) 0.367±0.133 7.25±0.19 Ophiuroidea…Глава 3. Стандартный обмен и критерий упорядоченности у позвоночных животных
Класс Amphioxi (Ланцетники)
Ланцетники, по мнению зоологов, являются наиболее древними животными в типе позвоночных. Исходя из этого, было бы желательным иметь как можно больше доброкачественных данных о стандартном обмене этих животных. К сожалению, это не так. Нам известны только четыре работы, в которых приводятся данные о стандартном обмене ланцетника Branchiostoma lanceolatum (Ивлев, 1961; Courtney, Newell, 1965; Ивлева, Попенкина, 1968, Ивлева, 1981). В трех последних работах приводятся сухая, но не сырая масса тела ланцетников, поэтому для расчета стандартного обмена мы использовали данные о содержании воды в теле ланцетников равной 78.5% (Виноградов, 1937). Расчет по данным указанных авторов показывает, что для Branchiostoma lanceolatum α=0.352, Cr=5.37 (Courtney, Newell, 1965), a=0.258, Cr=3.94 (Ивлева, Попенченко, 1969) и а=0.301, Сr=6.84 (Ивлева, 1981). С другой стороны, расчет по данным Ивлева (1961) показывает, что для этого вида ланцетника а=1.375, Сr=20.98. В целом без учета данных Ивлева имеем предварительные величины
КлассAmphioxia=0.304; Cr=5.38 (118)
Очевидно, что предстоит еще большая работа для того, чтобы получить правильные величины а и Сr для класса Amphioxi.
Рыбы
В состав этой группы в соответствии с принятой в настоящее время классификацией рыб (Решетников и др., 1989), мы включили четыре класса: Cephalaspidomorphi (Миноги), Myxini (Миксинообразные), Chondrichthyes (Хрящевые рыбы) и Osteichthyes (Костные рыбы).
Основная трудность изучения стандартного обмена у рыб, помимо всего прочего, связана с тем, что разные рыбы обладают разной степенью подвижности и при измерении скорости потребления кислорода рыбами исследователи сталкиваются с разной величиной вклада активного обмена в получаемую величину стандартного обмена. Избежать этой неточности очень трудно, поэтому во многих случаях мы имеем дело не со стандартным, а активным обменом, что приводит к завышению величины коэффициента а. Чтобы снизить величину ошибки при определении этого коэффициента можно было бы для активно плавающих рыб ввести поправку на активный обмен. Для этого можно было бы определять у рыб не стандартный, а "основной" обмен - обмен у неподвижных или наркотизированных рыб (Винберг, 1956). Оказалось, что чем больше подвижность рыб во время опыта, тем больше отличие стандартного обмена у нормальных и наркотизированных рыб: у разных видов рыб в зависимости от их подвижности "основной" обмен составляет от 40 до 100% от стандартного (Белокопытин, 1973). Следовательно, для внесения поправки необходимо иметь данные о дыхании неподвижных или наркотизированных рыб. Таких данных в литературе слишком мало, поэтому мы не можем учитывать это обстоятельство в своих расчетах, что, конечно, делает весьма условным приводимую нами классификацию рыб по стандартному обмену.
Следующая неточность связана с отсутствием достоверных данных о содержании воды в теле рыб. В сводке Виноградова (1937) приводятся многочисленные измерения содержания воды в теле рыб, но речь идет только о мягких частях тела. Поэтому при вычислении критерия упорядоченности у рыб,
как и у других позвоночных животных, мы приняли, содержание воды в теле равным 70%.
Данных о стандартном обмене рыб накопилось в литературе довольно много. Они обобщены в монографии Винберга (1956), где приведены данные о массе тела и скорости потребления кислорода 106 видов рыб (n=350) и в статье Озернюка и Булгаковой (1997) - 178 видов рыб (n=232). К сожалению монография Г.Г.Винберга написана слишком давно и в ней, естественно, не учтены работы последних 40 лет. В статье Озернюка и Булгаковой отражены более новые работы, но в ней не приведены данные о массе тела рыб, что не позволило нам пересчитать приводимые в этой работе величины коэффициента а с помощью принятого в данной монографии коэффициента k=0.75 (авторы приняли для своих расчетов k=0.87). Нам пришлось поэтому самим собрать данные о стандартном обмене рыб. Всего нами использованы данные о стандартном обмене 236 видов рыб (n=475).
При расположении и наименовании разных таксонов рыб мы использовали работы Расса (1971), а также Решетникова и др.(1989).
Классы Myxini (Миксинообразные) и Cephalaspidomorphi (Миноги).
Согласно расчетам Винберга (1956) аллометрическая зависимость скорости потребления кислорода от массы тела при 20°С у круглоротых (N=4), так же, как и костных рыб, имеет вид
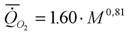
(здесь, как и везде мы выражаем коэффициент а в мВт).
Озернюк и Булгакова (1997) на основании данных о стандартном обмене 10 видов круглоротых получили несколько иные коэффициенты в аллометрической зависимости:
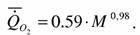
Значения аллометрических коэффициентов, приводимые Винбергом для круглоротых, по-видимому, имеют большую статистическую погрешность, так как при их вычислении автор использовал данные о стандартном обмене всего 4 видов миног (n=6). Озернюк и Булгакова также нашли в литературе
недостаточное число данных (3 вида миксин и 7 видов миног). Кроме того, при расчете коэффициентов в аллометрической зависимости Озернюк и Булгакова использовали не только данные о стандартном обмене взрослых миног, но и их личинок, что нельзя делать, так как пескорейки имеют значительно более низкий обмен, чем взрослые формы [табл. 34(1)].
Найденные нами материалы по дыханию круглоротых представлены в табл.34 (1). Расчеты по этим данным дают следующие значения аллометрических коэффициентов :
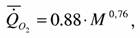
где k=0.764+0.113 достоверно не отличается от стандартной величины k=0.75. Поэтому, при расчетах коэффициента а мы использовали k=0.75.
При вычислении критерия упорядоченности мы считали, что содержание воды в теле круглоротых, как и у других позвоночных животных, близко 70%. С учетом сказанного имеем для круглоротых:
КлассMyxinia=0.237±0.094; Сr=4.23±1.68 (119)
Класс Cephalaspidomorphi a=1.025±0.128; Cr=15.27±1.91 (120)
взрослые формы a=1.440±0.095; Cr=21,45±1.42
Таблица 34(1)
Масса тела М(г), скорость потребления кислорода Q (мл О2/ч) при температуре t°C, сопоставимый стандартный обмен (коэффициент а, мВт) при 20 С и критерий упорядоченности (Cr) y рыб (классы: Myxini- Миксинообразные и Cephalaspidomorphi -Миноги).
| Классы и виды | M | Q | t | Автор | а | Cr |
| Myxini - Миксинообразные | 0.237 | 4.23 | ||||
| Eptatretus deani | 0.20 | 3.5 | (1) | 0.143 | 3.53 | |
| Eptatretus stoutii | 61.83 | 0.56 | (2) | 0.331 | 4.93 | |
| Cephalaspidomprphi Миноги взрослые формы | 1.025±0.128 1.440±0.095 | 15.27±1.91 21.45±1.42 | ||||
| Entosphenus tridentatus | 13.84 | (3) | 1.325 | 19.74 |
| Ichthyomyzon fossor | 3.78 | 0.40 | (4) | 0.992 | 14.78 | |
| Ichthyomyzon hubbsi | 0.156 | 15.5 | (5) | 0.455 | 6.78 | |
| Lampetra planeri | 5.2 | 0.79 | (6) | 1.286 | 19.17 | |
| 1.34 | 0.067 | (7) | 0.461 | 6.87 | ||
| 2.08 | 0.126 | (8) | 0.951 | 14.17 | ||
| Lampetra fluviatilis | 3.52 | 16.3 | (9) | 1.800 | 26.82 | |
| 1.27 | 0.065 | (8) | 0.710 | 10.57 | ||
| 2.50 | 12.7 | (10) | 1.198 | 17.85 | ||
| Lampetra japonica | 0.105 | (11) | 0.410 | 6.10 | ||
| Petromyzon marinus | 4.88 | 0.63 | (4) | 1.273 | 18.97 | |
| 75.6 | 6.56 | (11) | 1.435 | 21.39 |
(1) Smith,Hessler, 1974; (2) Munz,Morris, 1965; (3) Johansen et al., 1973; (4) Leach, 1946; (5) Hill,Potter, 1970; (6) Ивлев, 1954; (7) Potter,Rogers, 1972; (8) Lewis,Potter, 1977; Щербаков, 1937; (10) Claridge,Potter, 1975; (11) Beamish, 1973.
Класс Chondrichthyes(Хрящевые рыбы)
В обзоре Озернюка и Булгаковой (1997) использованы данные о стандартном обмене 9 видов акуловых рыб (п=9). Принималось, что аллометрическая зависимость дыхания от массы тела у акул такая же, как и у остальных рыб, т.е. k=0.87. Исходя из этого, авторы получили для этой группы рыб а=0.215 мл О2/г•час, т.е. в нашем обозначении а=1.206 мВт.
Расчеты по собранным нами данным дают следующую аллометрическую зависимость для хрящевых рыб:
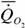 = 1.63·М0.76,
= 1.63·М0.76,
где k=0.763+0.054 достоверно не отличается от стандартной величины k=0.75. Поэтому, при расчетах коэффициента а мы использовали k=0.75.
В сводке Виноградова (1937) приводятся данные о содержании воды в мягких тканях хрящевых рыб. Оно равно в среднем 78.5% (N=13, n=19). Так как не совсем ясно, каково содержание воды в теле акул, включая скелет, то при расчете критерия упорядоченности мы приняли, что содержание воды в теле хрящевых рыб равно 70%. Собранные нами литературные данные о стандартном
обмене хрящевых рыб и рассчитанные величины коэффициента а (принимая, что k=0.75) и критерия упорядоченности Сr показаны в табл.34(2).
Мы использовали данные о стандартном обмене 9 видов акуловых рыб (n=16) [табл. 34(2)]. При расчете критерия упорядоченности было учтено, что многие хрящевые рыбы живут в субтропической или тропической зоне океанов. Имеем:
Класс Chondrichthyesa=1.766; Сr=27.40 (121)
Таблица 34(2)
Масса тела М(г), скорость потребления кислорода Q (мл О2/ч) при температуре t°C, сопоставимый стандартный обмен (коэффициент а, мВт) при 20°С и критерий упорядоченности (Сr) у рыб (класс Chondrichthyes- Хрящевые рыбы).
| Таксон | M | Q | t | Автор | a | Cr |
| Класс Chondrichthyes | 1.766±0.099 | 27.40±1.54 | ||||
| Подкласс Elasmobranchii | 1.766±0.099 | 27.40±1.54 | ||||
| Отряд Carcharhiniformes | 1.807±0.091 | 26.54±1.34 | ||||
| Scyliorhinus cassicula | 9.62 | (1) | 1.510 | 22.50 | ||
| Scyliorhinus canicula | 8.94 | (1) | 1.957 | 29.16 | ||
| 23.7 | (2) | 1.727 | 25.80 | |||
| 8.785 | 12.2 | (3) | 2.009 | 29.93 | ||
| Scyliorhinus stellaris | 59.95 | (1) | 1.949 | 28.80 | ||
| 107.64 | (4) | 2.343 | 34.91 | |||
| 39. 19 | 18.5 | (5) | 1.886 | 28.10 | ||
| 83.71 | (6) | 2.064 | 30.76 | |||
| Scyllium cassicula | 132.6 | 4.96 | (5) | 1.311 | 19.24 | |
| Scyllium catulus | 16.78 | (5) | 1.497 | 22.30 | ||
| 21.5 | 1.776 | (5) | 1.651 | 24.73 | ||
| Отряд Orectolobiformes | 1.172 | 28.99 | ||||
| Hemiscyllium plagiosum | (7) | 1.172 | 28.99 | |||
| Отряд Squaliformes | 2.195 | 32.71 | ||||
| Squalus suckleyi | 14.91 | (8) | 2.218 | 33.05 | ||
| (1) | 2.172 | 32.36 | ||||
| Отряд Rajiformes | 1.842 | 27.68 | ||||
| Raja torpedo | 17.14 | 14.5 | (5) | 1.842 | 27.68 | |
| Отряд Torpediniformes | 0.950 | 23.49 | ||||
| Torpedo marmorata | 29.3 | (1) | 0.950 | 23.49 |
(1) Hughes, 1978; (2) Butler,Taylor, 1975; (3) Hughes,Umezawa, 1968; (4) Piiper et al., 1970; (5) Винберг, 1956; (6) Baumgarten-Schumann,Piiper, 1968; (7) Chan,Wong, 1977; (8) Gerald, Cech, 1970.
Класс Osteichthyes(Костные рыбы)
Озернюк и Булгакова (1997) приводят данные о дыхании 159 видов (n=210) костных рыб. Аллометрическая зависимость стандартного обмена от массы тела (этих и акуловых рыб) по расчетам этих авторов имеет вид:
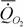 = 0.98М0.87.
= 0.98М0.87.
Мы собрали данные о стандартном обмене 228 видов рыб (n=446) и получили аллометрическую зависимость вида
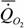 = 1.26М0.84,
= 1.26М0.84,
где k=0.844±0.080. Другими словами, коэффициент k достоверно не отличается от k=0.75.
Таблица 34(3)
Сопоставимый стандартный обмен (коэффициент а, мВт) при 20°С и критерий упорядоченности (Сr) у рыб (класс Osteichthyes- Костные рыбы). N - число изученных видов, n - число измерений.
| Таксон | а | Сr | N | n |
| Подкласс Sarcopterygii | 0.570±0.163 | 14.10±4.03 | ||
| Надотряд Dipnoi (Двоякодышащие) | 0.570±0.163 | 14.10±4.03 | ||
| Отряд Ceratodontiformes | 0.327 | 8.10 | ||
| Отряд Lepidosireniformes | 0.732 | 18.10 | ||
| Подкласс Actinopterygii | 1.634±0.231 | 24.21±3.42 | ||
| Надотряд Ganoidomorpha | 1.634±0.231 | 24.21±3.42 | ||
| Отряд Polypteriformes | 0.653 | 16.15 | ||
| Отряд Lepisosteiformes | 0.984 | 19.49 | ||
| Отряд Acipenseriformes | 1.974±0.211 | 29.42±3.14 | ||
| Подкласс Teleostei | 1.816±0.047 | 30.46±0.79 | ||
| Надотряд Batrachomorpha | 0.301 | 7.44 | ||
| Надотряд Anguillomorpha | 1.208±0.148 | 24.98±3.06 | ||
| Отряд Anguilliformes | 1.208±0.148 | 24.98±3.06 | ||
| Сем. Morhguidae | 0.306 | 7.57 | ||
| Сем. Anguillidae | 1.107±0.054 | 21.02±1.03 | ||
| Сем. Congridae | 1.715 | 42.42 | ||
| Сем. Muraenidae | 2.610 | 64.55 |
| Надотряд Cyprinomorpha | 1.639±0.067 | 25.34±1.04 | ||
| Отряд Cypriniformes | 1.774±0.071 | 26.85±1.07 | ||
| Подотряд Gymnotoidei | 0.537 | 13.28 | ||
| Сем. Gymnotidae | 0.347 | 8.59 | ||
| Сем. Electrophoridae | 0.726 | 17.96 | ||
| Подотряд Cyprinoidei | 1.808±0.071 | 27.10±1.06 | ||
| Сем. Cobitidae | 1.283±0.191 | 19.12±2.85 | ||
| Сем. Catostomidae | 1.413 | 21.05 | ||
| Сем. Cyprinidae | 1.860±0.076 | 27.89±1.14 | ||
| Отряд Siluriformes | 0.877±0.053 | 16.80±1.02 | ||
| Сем. Heteropneustidae | 0.536 | 13.26 | ||
| Сем. Ictaluridae | 0.950±0.047 | 14.16±0.70 | ||
| Сем. Loricariidae | 0.638 | 15.79 | ||
| Сем. Plotosidae | 0.651 | 16.11 | ||
| Сем. Bagridae | 0.796 | 19.68 | ||
| Сем. Saccobranchidae | 0.804 | 19.88 | ||
| Сем. Callichthyidae | 1.023 | 25.32 | ||
| Сем. Clariidae | 1.094 | 27.66 | ||
| Надотряд Atherinomorpha | 1.492±0.198 | 32.58±4.32 | ||
| Отряд Cyprinodontiformes | 1.492±0.198 | 32.58±4.32 | ||
| Сем. Poeciliidae | 1.317±0.326 | 28.00±6.93 | ||
| Сем. Cyprinodontidae | 1.461±0.119 | 36.14±2.94 | ||
| Сем. Oryziatidae | 2.903 | 43.26 | ||
| Надотряд Percomorpha | 1.837±0.080 | 32.59±1.42 | ||
| Отряд Scorpaeniformes | 1.652±0.181 | 29.27±3.21 | ||
| Подотряд Cottoidei | 1.529±0.211 | 26.51±3.66 | ||
| Сем. Liparidae | 0.800 | 11.92 | ||
| Сем. Cottidae | 1.727±0.235 | 30.49±4.15 | ||
| Подотряд Scorpaenoidei | 1.898±0.345 | 34.79±6.32 | ||
| Сем. Scorpaenidae | 1.267 | 25.11 | ||
| Сем. Triglidae | 2.739 | 47.70 | ||
| Отряд Pleuronectiformes | 1.762±0.182 | 29.49±3.05 | ||
| Подотряд Pleuronectoidei | 1.777±0.199 | 29.11±3.26 | ||
| Подотряд Soleoidei | 1.691 | 31.27 | ||
| Сем. Cynoglossidae | 0.648 | 16.04 | ||
| Сем. Soleidae | 2.212 | 38.89 | ||
| Отряд Mugiliformes | 1.411 | 29.95 | ||
| Отряд Gasterosteiformes | 1.739±0.233 | 31.60±4.23 | ||
| Подотряд Syngnathoidei | 1.480±0.224 | 28.87±4.37 | ||
| Сем. Syngnathidae | 1.437±0.220 | 27.55±4.22 | ||
| Сем. Carangidae | 1.671 | 34.79 | ||
| Подотряд Gasterosteoidei | 2.311 | 37.62 | ||
| Отряд Perciformes | 1.884±0.106 | 33.09±1.86 | ||
| Подотряд Zoarcoidei | 1.115±0.243 | 16.61±3.62 | ||
| Подотряд Anabantoidei | 0.862 | 21.31 | ||
| Сем. Anabanyidae | 0.831 | 20.55 | ||
| Сем. Chanidae | 0.892 | 22.07 |
| Подотряд Trachinoidei | 1.733 | 25.82 | ||
| Подотряд Gobioidei | 1.305±0.194 | 27.30±4.06 | ||
| Сем. Gobiidae | 1.148±0.167 | 25.00±3.34 | ||
| Сем. Periophthalidae | 2.324 | 42.26 | ||
| Подотряд Callionymoidei | 1.914 | 28.51 | ||
| Подотряд Percoidei | 1.742±0.210 | 31.73±3.83 | ||
| Сем. Cepolidae | 0.326 | 6.33 | ||
| Сем. Grammistidae | 0.496 | 12.27 | ||
| Сем. Kuhliidae | 0.586 | 14.49 | ||
| Сем. Centrarchidae | 1.055±0.155 | 17.95±2.64 | ||
| Сем. Cichlidae | 0.934±0.091 | 23.11 | ||
| Сем. Serranidae | 1.302 | 32.19 | ||
| Сем. Percidae | 2.232±0.176 | 33.47±2.06 | ||
| Сем. Centracanthidae | 1.705±0.195 | 33.78±3.86 | ||
| Сем. Embiotocidae | 2.289 | 34.11 | ||
| Сем. Mullidae | 1.643 | 40.63 | ||
| Сем. Girellidae | 1.742 | 43.08 | ||
| Сем. Percichthydae | 3.424±0.375 | 51.02±5.59 | ||
| Сем. Pomadagyidae | 2.087 | 51.62 | ||
| Сем. Sparidae | 2.891±0.490 | 58.31±9.88 | ||
| Сем. Sciaenidae | 3.033 | 75.02 | ||
| Подотряд Labroidei | 1.423 | 31.94 | ||
| Подотряд Blennioidei | 1.705 | 35.89 | ||
| Подотряд Notothenioidei | 3.443±0.372 | 51.31±5.54 | ||
| Сем. Bathydraconidae | 2.490 | 37.10 | ||
| Сем. Chaenichthyidae | 2.748±0.477 | 40.94±7.11 | ||
| Сем. Nototheniidae | 3.846±0.504 | 57.31±7.51 | ||
| Подотряд Echeneoidei | 2.885 | 71.35 | ||
| Надотряд Parapercomorpha | 2.114±0.336 | 33.00±5.25 | ||
| Отряд Gadiformes | 2.114±0.336 | 33.00±5.25 | ||
| Подотряд Ophidoidei | 0.302 | 7.46 | ||
| Подотряд Gadoidei | 2.316±0.300 | 35.83±4.64 | ||
| Сем. Macrouridae | 0.359 | 8.87 | ||
| Сем. Gadidae | 2.560±0.198 | 39.20±5.48 | ||
| Надотряд Osteoglossomorpha | 1.416 | 35.03 | ||
| Надотряд Clupeomorpha | 2.378±0.090 | 35.79±1.35 | ||
| Отряд Salmoniformes | 2.331±0.082 | 34.72±1.22 | ||
| Подотряд Osmeroidei | 1.813 | 27.01 | ||
| Подотряд Esocoidei | 2.011±0.194 | 29.97±2.89 | ||
| Подотряд Salmonoidei | 2.390±0.089 | 35.27±1.31 | ||
| Сем. Salmonidae | 2.313±0.089 | 34.46±1.33 | ||
| Сем. Coregonidae | 2.803±0.324 | 41.77±4.83 | ||
| Сем. Thymallidae | 3.424 | 51.01 | ||
| Отряд Gonorhynchiformes | 2.131 | 52.70 | ||
| Отряд Clupeiformes | 3.779 | 56.31 |
Рассчитанные по собранным нами литературным данным (Зотин, Зотин, 1999) коэффициенты а и Сr (при k=0.75) представлены в табл. 34(3). Классификация костных рыб в этой таблице дана по Рассу (1971) и Решетникову и др. (1989), но расположение отдельных таксонов приведено в соответствии с величиной критерия упорядоченности. Расположение отрядов в табл. 34(3) отличается от приводимого Озернюком и Булгаковой (1997), хотя принципы заложенные в основу этой и нашей работы почти одинаковые. Это показывает насколько еще ненадежно, при имеющихся в настоящее время в литературе данных о стандартном обмене рыб, использование коэффициента а и критерия упорядоченности для построения макросистематики рыб. И все же расположение подклассов в классе Osteichthyes можно представить согласно данным, приводимым в табл.34 (3) в следующем виде:
Sarcopterygii -> Actinopterygii -> Teleostei,
а предварительное расположение надотрядов в подклассе Teleostei в виде:
Batrachomorpha -> Anguillomorpha -> Cyprinomorpha -> —> Atherinomorpha —> Percomorpha —> Parapercomorpha —> -> Osteoglossomorpha-> Clupeomorpha
Следует отметить, что, по нашему мнению, подклассы Sarcopterygii, Actinopterygii и Teleostei следовало бы выделить в отдельные классы. Мы пока этого не делаем, но запишем величину коэффициента а (при k=0.75) и критерия упорядоченности отдельно для каждого из указанных подклассов. В результате мы имеем
Класс Osteichthyes a=1.799; Cr=30.14 (122)
Подкласс Sarcopterygii a=0.570; Cr=14.10
Подкласс Actinopterygii a=1.455; Cr=24.21
Подкласс Teleostei a=1.820; Cr=30.46
Класс Amphibia (Земноводные)
В работах И.Г.Владимировой и др. (Владимирова, Сербинова, 1992; Зотин, Владимирова, 1993; Владимирова, Зотин, 1994) приведены экспериментальные и литературные данные о массе тела и интенсивности потребления кислорода у представителей класса Amphibia: 4 вида из отряда Gymnophiona (n=4), 69 видов из отряда Caudata (n=149) и 85 видов из отряда Anura (n=208). Для сравнения стандартного обмена разных видов амфибий данные были пересчитаны по формуле зависимости дыхания от температуры вида (68):
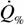 = 100•exp[0.0875•(t0 - 20)],
= 100•exp[0.0875•(t0 - 20)],
если авторы проводили свои измерения не при 20°С.
Позднее Владимирова и др. (Vladimirova et al., 1997) получили дополнительные данные о 8 видах амфибий и особенно таких примитивных хвостатых амфибиях, как углозуб (Salamandrella keyserlingii) и лягушкозуб (Ranodon sibiricus) из семейства Hynobiidae. На основании этих и собранных ранее (Владимирова, Зотин, 1994) данных на рис.24 в логарифмической шкале координат сопоставлены скорость потребления кислорода и масса тела у хвостатых и бесхвостых земноводных. Полученные прямые заметно отличаются у хвостатых и бесхвостых амфибий, но в обоих случаях зависимость дыхания от массы тела: во-первых, соответствует аллометрическому соотношению вида (46), во-вторых, параллельны друг другу, т.е. имеют сходный коэффициент k. Расчет методом, описанным ранее (Зотин, 1999), показал, что для всего класса амфибий в целом при 20°С k =0.764±0.052. Отличные, а иногда близкие, величины получили другие авторы: k=0,817 при 15°С для 29 видов хвостатых (Feder, 1976); k=0,809 при 15°С для 26 видов бесхвостых (Mac Nally, 1981); k=0,856 при 15°С для 7 видов легочных и k=0,72 для 4 видов безлегочных саламандр (Whitford, Hutchison, 1967); k=0.19 при 25°С (Bucher et al., 1982) и k=0.13 при 20° для 17 видов бесхвостых амфибий (Taigen, 1983). В работе Владимировой и Зотина (1994) показано, что для хвостатых и бесхвостых амфибий k=0.81.
Так как в настоящей работе мы использовали более корректный статистический метод (Зотин, 1999), чем в вышеуказанных работах, то величина k=0,76 должна быть ближе к реальности и к стандартной величине k=0.75,
которую мы и использовали для расчетов коэффициента а и критерия упорядоченности Сr для разных видов, семейств, отрядов и всего класса амфибий в целом. Пересчет был произведен по данным, приведенным в обзоре Зотина и Владимировой (1993) и измерениям Владимировой и др. (Vladimirova et al., 1997) [табл.35].
Таблица 35.
Коэффициент а (мВт/г) при 20° С и критерий упорядоченности Сr в разных отрядах и семействах класса Amphibia(Земноводные). N - число изученных видов, n - число измерений.
| Таксон | а | Сr | N | n |
| Отряд Caudata | 0.394±0.014 | 7.84±0.34 | ||
| Сем. Hynobiidae | 0.368 | 5.49 | ||
| Сем. Amphiumidae | 0.396±0.087 | 5.90±1.30 | ||
| Сем. Plethodontidae | 0.351±0.013 | 6.51±0.27 | ||
| Сем. Cryptobranchidae | 0.486 | 7.24 | ||
| Сем. Sirenidae | 0.353±0.062 | 7.82±0.71 | ||
| Сем. Dicamptodontidae | 0.571 | 8.50 | ||
| Сем. Salamandridae | 0.415±0.032 | 8.65±0.81 | ||
| Сем. Proteidae | 0.414±0.067 | 8.86±1.15 | ||
| Сем. Ambystomatidae | 0.573±0.073 | 14.07±1.86 | ||
| Отряд Gymnophiona | 0.509+0.11 | 12.60±2.94 | ||
| Сем. Caeciliidae | 0.500 | 12.36 | ||
| Сем. Typhlonectidae | 0.524 | 12.96 | ||
| Отряд Anura | 0.966±0.029 | 16.49±0.48 | ||
| Сем. Discoglossidae | 0.582 | 9.88 | ||
| Сем. Microhylidae | 0.496 | 12.28 | ||
| Сем. Hyperoliidae | 0.506 | 12.51 | ||
| Сем. Rhacophoridae | 0.887 | 13.21 | ||
| Сем. Hylidae | 0.915±0.060 | 14.80±0.92 | ||
| Сем. Bufonidae | 1.045+0.080 | 15.57±1.31 | ||
| Сем. Leptodactylidae | 0.702±0.061 | 15.92±1.34 | ||
| Сем. Dendrobatidae | 0.657±0.056 | 16.26±1.37 | ||
| Сем. Ranidae | 1.083+0.046 | 16.55±0.72 |
| Сем. Pipidae | 0.976±0.094 | 23.12±2.09 | ||
| Сем. Pelobatidae | 1.180+0.16 | 25.72±4.51 |
Как видно из этих расчетов, стандартный обмен бесхвостых земноводных превышает стандартный обмен хвостатых более чем в 2.1 раза. В случае отряда безногих мы не можем говорить с такой же определенностью об отличиях представителей этого отряда от бесхвостых амфибий, так как в литературе имеется слишком мало данных о дыхании безногих. Но, судя по данным, приводимым в табл.35, средний коэффициент а для бесхвостых амфибий превышает величину этого коэффициента для безногих амфибий примерно в 1.3 раза.
При расчете критерия упорядоченности Сr учитывалось, что амфибии являются полуводными и водными животными. Для водных животных мы приняли в качестве эталона содержание воды в теле равным 86.5%, для наземных - 70%. Однако не исключено, что у всех позвоночных животных содержание воды в теле одинаково, поэтому мы предпочли при определении критерия упорядоченности для амфибий считать, что содержание воды в них соответствует стандарту 70%, как в случае наземных, так и водных животных.
Подсчет показал, что у наземных амфибий умеренного пояса коэффициенТабольше, чем тропического: в отряде хвостатых примерно в 1.56 раза, в отряде бесхвостых в 1.44 (Владимирова, Зотин, 1994). Эти цифры близки к величине аu /at=1.66, которую мы приняли в качестве стандарта для всех животных и будем использовать при расчете критерия упорядоченности наземных представителей класса амфибий. Для водных амфибий, однако, различий коэффициентов а в зависимости от широты обитания не наблюдается (Владимирова, Зотин, 1994). Учитывая сказанное, мы получили для класса амфибий несколько иные цифры критерия упорядоченности, чем в работе Владимировой и Зотина (1994).
Как видно из табл.35, в среднем критерий упорядоченности отряда хвостатых земноводных в 2,45 раза меньше, чем в отряде бесхвостых (у хвостатых - 5.87; у бесхвостых - 14.39). Это соответствует более примитивному строению хвостатых и более раннему появлению этого отряда в процессе
эволюции по сравнению с бесхвостыми (Кашкаров, Станчинский, 1940; Шмальгаузен, 1964; Наумов, Карташов, 1979; Halliday, Adler, 1986).
Судя по палеонтологическим данным отряд безногих земноводных появился значительно позже хвостатых и бесхвостых (Halliday, Adler, 1986). Однако пока получено слишком мало данных (4 вида, 4 измерения) для того, чтобы судить о том противоречит ли критерий упорядоченности безногих их более позднему возникновению: исходя из материалов, приводимых в табл.35, отряд Gymnophiona занимает промежуточное положение между Anura и Caudata, имея критерий упорядоченности порядка 12.60 против 16.49 у Anura.
В соответствии с увеличением критерия упорядоченности Сr можно расположить 3 отряда этого класса следующим образом (Владимирова, Зотин, 1994):
Caudata -> Gymnophiona -> Anura
Еще труднее из-за ограниченности данных о стандартном обмене построить по критерию упорядоченности макросистематику семейств в отдельных отрядах… Dicamptodontidae, для которых имеются только единичные наблюдения о стандартном обмене) не противоречит принятому некоторыми авторами делению этого отряда амфибий на…Testudintes —> Rhynchocephalia —> Squamata —> Crocodylia
Явное несоответствие выявляется и для семейства ящериц, представители которых известных с эоцена, т.е. с того же времени, когда возникли и другие… При определении критерия упорядоченности учитывалось, что большинство видов… Класс Reptilia a=0.861±0.027; Cr=18.22±0.49 (124)Gt; Chiroptera —> Primates —> Cetacea —> Perissodactyla
Конечно, представленное в табл.38 и на схеме расположение отрядов это только предварительная макросистема инфракласса Eutheria класса млекопитающих,… В целом для млекопитающих коэффициент а и критерий упорядоченности Сr равны: … Класс Mammaliaa=24.29±0.09; Cr=104.0±0.2 (125) Отряд Insectivora a=39.50±0.50; Cr=155.8±0.96Глава 4. Прогрессивная эволюция и биоэнергетический прогресс
Согласно термодинамике неравновесных процессов, при протекании необратимых процессов любая термодинамическая система стремится к равновесию, т.е. к наиболее вероятному состоянию. В то же время, согласно принципу наискорейшего спуска в достаточно сложной системе существуют
подсистемы, которые двигаются в обратном направлении от состояния равновесия, т.е. от более вероятного к менее вероятному состоянию. Это можно отнести и к такой сложной системе, как биосфера: в целом она движется к состоянию равновесия, но существует ряд таксонов организмов, которые эволюционируют в противоположном направлении. Изменение таких организмов мы и называем прогрессивной эволюцией.
Как отмечалось, эволюция любой открытой системы сопровождается изменением функции внешней диссипации: она уменьшается при приближении системы к равновесию и увеличивается при удалении от равновесия. Так как для живых организмов функцию внешней диссипации с известным приближением можно приравнять удельной скорости потребления кислорода организмами, т.е. стандартному обмену, то основным признаком прогрессивной эволюции является увеличение стандартного обмена, входящего в состав критерия упорядоченности. Хотя прогрессивная эволюция основана на изменении энергетического метаболизма, она не идентична биоэнергетическому прогрессу, так как прогрессивная эволюция предполагает не только усиление энергетики, но и совершенствование морфологического строения, поведения и функционирования организмов в процессе эволюции
Изменение энергетического обмена в ходе эволюции
Таблица41 Сводная таблица коэффициентов а (мВт) и критериев упорядоченности Сr в разных… Таксон а N n Сr Простейшие …Биологический и физический смысл эволюционного прогресса
Согласно Печуркину (1982) существует магистральное направление эволюции организмов, связанное со способностью захватывать и использовать потоки… Биоэнергетический прогресс. Биологический смысл направленности этого процесса… Максимальный обмен у всех животных независимо от размеров и систематического положения прочно связан с величиной…Направление и скорость прогрессивной эволюции и биоэнергетического прогресса
Исходя из данных, приводимых в табл..41 и рис.28, можно считать, что биоэнергетический прогресс и прогрессивная эволюция в животном царстве… Простейшие —> Гребневики —> Кишечнополостные —> —> Черви —>… в типе моллюсков:Направление и скорость эволюционного прогресса в соответствии с константой Рубнера
Ru = 6.81•ехр(0.010·tg) (144) Бауэр (1935) для своих выводов смог воспользоваться единичными измерениями… Однако при расчете константы Рубнера, где используются данные о максимальной продолжительности жизни в разных отрядах…Аллогенез и катагенез
Как отмечалось, согласно Северцову (1925, 1934, 1939) биологический прогресс, т.е. успех в борьбе за существование, связан не только с… Теперь имеет смысл разобрать вопрос о том, как соотносится с теорией Северцова… В соответствии с теорией Северцова биологический прогресс выражается не только в арогенезе, но и аллогенезе, когда…Часть 3. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: СОВЕРШЕНСТВОВАНИЕ РЕГУЛЯЦИИ И УПРАВЛЕНИЯ
"Знаю, что едва ли возможно определить Ясно, что разумеется под более высокой Или более низкой организацией. ЭтоКритерии организованности
В живых системах, которые можно определить, как открытые, далекие от равновесия, организованные системы, способные к саморазмножению и саморазвитию,… Кроме того, в живых системах протекает множество эндотермических, сопряженных… Из указанных негэнтропийных эффектов реально измеримым в настоящее время является только интегральный негэнтропийный…Глава 1. Температурный гомеостаз
Как отмечалось, в настоящее время для количественной характеристики совершенства регуляции в организмах реально можно использовать критерии надежности гомеостазов (150, 151). Трудность состоит в том, что параметры, входящие в формулы (150) и (151) должны иметь четкий физический смысл и быть достаточно легко измеряемыми. К сожалению пока такого рода трудность преодолена только в отношении температурного гомеостаза у гомойотермных животных (Зотин, 1988; Владимирова, Зотин, 1989) и, частично, в отношении гомеореза пойкилотермных животных (Alekseeva et al., 1992; Зотин, Зотина, 1993; Zotin, 1995). Ниже мы остановимся только на проблемах, связанных с температурным гомеостазом, так как критерии, характеризующие гомеорез, получены пока в ограниченном числе.
Гомеостаз, поддерживающий постоянство температуры тела гомойотермных животных, имеет свои особенности, которые следует учитывать при вычислении меры надежности и зоны оптимальности. Кривая терморегуляции, которая показывает, сколько энергии затрачивается на поддержание температуры тела приблизительно на постоянном уровне, носит довольно сложный характер (рис.10).
Одной из особенностей этой кривой является то, что в области химической терморегуляции потребление кислорода практически линейно зависит от температуры среды:
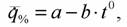 (153)
(153)
поэтому для вычисления метаболитической цены химической терморегуляции можно использовать несколько видоизмененную формулу Хаскина (1975)
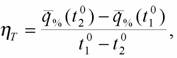 (154)
(154)
где ηT - относительная метаболитическая цена химической терморегуляции; 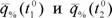 - интенсивность потребления кислорода животным выраженная в процентах от потребления кислорода в ТНЗ, принятой за 100%; t10 и t20 - температура среды.
- интенсивность потребления кислорода животным выраженная в процентах от потребления кислорода в ТНЗ, принятой за 100%; t10 и t20 - температура среды.
Очевидно, что чем меньше затрачивается энергии на поддержание температуры тела при изменении ее в окружающей среде, тем надежней и совершенней терморегуляция организма. Поэтому метаболитическая цена
химической терморегуляции может служить критерием совершенства температурного гомеостаза. Следует учитывать при этом, что интенсивность потребления кислорода в оптимальной точке (стандартный обмен) значительно отличается у разных видов гомойотермных животных. Поэтому в формуле (147) метаболитическая цена выражена в относительных величинах (Зотин, 1988; Владимирова, Зотин, 1989). Соответственно, мера надежности терморегуляции для гомойотермных животных равна:
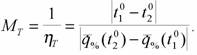 (155)
(155)
Критерий (155) введен, опираясь на линейную аппроксимацию кривой химической терморегуляции (153). Но, как видно из схемы, показанной на рис.10, кривая терморегуляции в своей нижней части имеет вид параболы и, поэтому, правильней было бы ее описывать уравнением параболы (Studier, O'Farrell, 1976). Однако и такое более сложное уравнение, не может полностью аппроксимировать кривую терморегуляции и особенно максимум в зоне низких температур (рис.10). Можно, однако, упростить задачу, используя, с одной стороны, уравнение (149) при описании нижней части кривой терморегуляции, с другой стороны, уравнение (153), для описания области химической терморегуляции. Это оправдывается тем, что уравнение параболы (149) позволяет ввести второй критерий надежности терморегуляции - зону оптимальности температурного гомеостаза или, как ее принято называть, термонейтральную зону (ТНЗ), которая может быть вычислена, согласно (151) по формуле
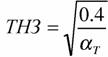 (156)
(156)
где αT - констант из уравнения параболы для терморегуляции вида (149). Уравнение параболы (148) позволяет также определить оптимальную (или, как ее иногда называют, критическую) точку (рT) кривой терморегуляции (рис.10) по формуле
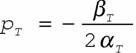 (157)
(157)
Соотношения (155 - 157), в принципе, можно использовать для сравнения совершенства температурного гомеостаза в разных группах гомойотермных животных. Следует, однако, учитывать одну особенность температурного гомеостаза, которая, возможно, свойственна и некоторым другим конкретным видам гомеостазов. Речь идет о том, что мера надежности терморегуляции (155), термонейтральная зона (156) и оптимальная точка (157) зависят от массы тела животного (Зотин, 1988; Владимирова, Зотин, 1989). Сопоставление величины меры надежности химической терморегуляции с массой тела гомойотермных животных позволило получить поправки, связанные с различием в размерах тела животных. В этом случае сопоставимую меру надежности температурного гомеостаза (МT ) для млекопитающих можно записать в виде:
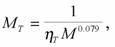 (158)
(158)
а для птиц
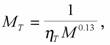 (159)
(159)
где МT - сопоставимая величина меры надежности температурного гомеостаза, т.е. мера надежности гомойотермных животных с условной массой в 1 г.
Зависимость ширины ТНЗ от массы тела получена пока только для млекопитающих и условная зона оптимальности для организма массой в 1 г (ΖT ) у этих животных равна (Зотин, 1988; Владимирова, Зотин, 1989):
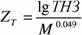 (160)
(160)
Поскольку мера надежности МT и зона оптимальности zt взаимосвязаны (152), мы в дальнейшем будем приводить только величину меры надежности.
Совершенствование терморегуляции в процессе прогрессивной эволюции
эволюционной лестнице". Эта идея была проверена (Владимирова, Зотин, 1989; Зотин и др., 1990). с использованием данных о кривых терморегуляции… Таблица 46 Сопоставимая мера надежности (МT) у представителей основных отрядов млекопитающих (Зотин, Владимирова, 1991).Адаптация к условиям существования.
что и было сделано (Владимирова, Зотин, 1989). Проведенные этими авторами расчеты для нескольких отрядов млекопитающих представлены в табл.47. Как… Таблица 47 Основные показатели, характеризующие кривые терморегуляции у млекопитающих, живущих в разных климатических зонах: МT -…Скорость совершенствования терморегуляции у млекопитающих в процессе эволюции.
Мы уже отмечали, что хотя энергетические показатели имеют хорошую теоретическую основу, они недостаточны для построения макросистематики животных. В… С учетом сделанных замечаний мы можем расположить отряды в макросистеме класса… Monotremata (Tachyglossidae) -> Marsupialia -> Edentata -> -> Chiroptera -> Insectivora ->…Gt; Lagomorpha -> Perissodactyla
Расположение отрядов по критериям температурного гомеостаза показывает, что прогрессивная эволюция млекопитающих сопровождалась совершенствованием… На основе данных, приводимых в табл.46 и 42 можно подсчитать скорость эволюции… МT = 0.018exp(0.0094tg) (162)Суперорганизмы.
У разных суперорганизмов терморегуляция осуществляется разными способами. В муравейниках - за счет целесообразной деятельности муравьев, в рое пчел… Хотя отдельные муравьи (как пойкилотермные организмы) не способны к… открывают входы, способствующие проветриванию муравейника и снижению температуры в ядре. При охлаждении муравейника…Глава 2. Максимальная продолжительность жизни
Изменение организованности животных и растений в процессе прогрессивной эволюции можно характеризовать, как говорилось, различными количественными критериями. Одним из таких критериев является величина максимальной продолжительности жизни. Дело в том, что длительность жизни животных и растений, как и искусственных приборов и аппаратов, несомненно связана с совершенством и надежностью их системы регуляции и управления, кроме может быть тех случаев когда вид заинтересован в преждевременной смерти его представителей (гибель кеты после нереста, отсутствие ротового аппарата у самцов некоторых насекомых и т.д.).
К сожалению, точное определение максимальной продолжительности жизни большинства животных не всегда возможно (кроме может быть человека и некоторых домашних животных). Это связано с тем, что у диких животных, живущих на свободе, максимальная продолжительность жизни за редким исключением неосуществима, так как стареющие особи менее приспособлены к борьбе за существование и погибают раньше положенного срока. Например, судя по данным, приводимым Паевским (1985а), у птиц, живущих в неволе, максимальная продолжительность жизни на 48% (n=41) больше, чем у птиц, живущих на воле. В свою очередь дикие животные, живущие в неволе, постоянно находятся в стрессовых условиях и не ясно насколько правильно они содержатся. То, что условия содержания значительно влияют на продолжительность жизни в зоопарках ясно из сравнения максимальной продолжительность жизни млекопитающих, приводимой в списке Флауэра (Flawer, 1931) и Джонеса (Jones, 1982): за пятьдесят лет улучшения условий жизни в зоопарках максимальная продолжительность жизни животных заметно возросла.
Прежде чем переходить к характеристике отдельных групп животных по этому признаку следует напомнить, что максимальная продолжительность жизни, как и остальные признаки, находится в аллометрической зависимости от массы тела животных (Ordy, 1976; Economes, 1980; Calder, 1983, 1984; Prinzinger, 1989):
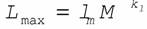 (163)
(163)
где M - масса животного в кг; Lmax - максимальная продолжительность жизни в годах; 1m - сопоставимая максимальная продолжительность жизни (продолжительность жизни для данного вида животного, если бы их масса равнялась 1 кг).
На рис.36 представлены данные о максимальной продолжительности жизни и массе тела животных, взятых из различных обзоров и монографий (Flower, 1931; Нагорный и др., 1963; Comfort, 1979; Zepelin, Rechtschaffen, 1974; Outer, 1978; Tolmasoff et al., 1980; Jones, 1982; Паевский, 1985а; Jürgens, Prothero, 1987, 1991; Гаврилов, 1991; Зотин, 1993а,б). В логарифмической шкале координат имеет место линейная зависимость между этими показателями. Расчет методом наименьших квадратов дал следующие величины коэффициента: для приматов k1=0.26 (Ordy, 1976); для всех млекопитающих k1=0.17 (Economes, 1980); k1=0.16 (Calder, 1983); k1=0.17 (Jürgens, Prothero, 1991); k1=0.23 (Зотин, 1993a); для птиц k1=0.20 (Lindstedt, Calder, 1976); k1=0.18 (Calder, 1983); k1=0.15 (Паевский, 19856); k1=0.23 (Prinzinger, 1989); для воробьиных птиц k1=0.23 (Calder, 1974); k1=0.26 (Lindstedt, Calder, 1976); для куликов k1=0.24 (Гаврилов, 1991). Использование статистически более правильного метода (А.А.Зотин, 1998) показало, что для млекопитающих k1= 0.122 + 0.021, а для птиц - k1=0.16 ±0.03. Аллометрические зависимости максимальной продолжительности жизни от массы животных равны:
для млекопитающих Lmax = 5.24· Μ0.14, (164)
для птиц Lmax = 7.17 · M0·17 (165)
В дальнейшем мы будем использовать величины 1т и Lmax из формулы (163) при расчетах величины максимальной продолжительности жизни в отдельных таксонах млекопитающих и птиц. Чтобы не осложнять себе жизнь мы ограничимся в основном только этими классами типа позвоночных животных.
Казалось бы, что и в этом случае мы должны сравнивать различных животных не по абсолютной величине продолжительности жизни Lmax, а по
коэффициенту 1^ из аллометрического соотношения (163). Однако следует учитывать, во-первых, средняя продолжительность данного вида животных (тем более, максимальная) сильно зависит от внутри- и межвидовой конкуренции: чем выше давление отбора, тем меньше средняя и максимальная продолжительность жизни; во-вторых, максимальная продолжительность жизни зависит не только от массы тела животного, но в еще большей степени от относительных размеров мозга. На основании сопоставления продолжительности жизни и коэффициента энцефализации у 400 видов млекопитающих показано (Зотин, 1993 б), что между этими показателями имеет место линейная зависимость (рис.37). Линейная зависимость между коэффициентом энцефализации и продолжительностью жизни наблюдается не только у млекопитающих, но и у других позвоночных животных, хотя наклон прямых в разных классах различен (Mallouk, 1975; Martin, 1981). По мнению Маллоука (Mallouk, 1975) корреляция между максимальной продолжительностью жизни и относительными размерами мозга значительно более тесная, чем между любыми другими зависимостями для продолжительности жизни животных. На этом основании он высказал предположение об особом «жизненном веществе», которое синтезируется мозгом во время его роста, а затем только расходуется в процессе жизни, что в конце концов и приводит к естественной смерти.
Гипотеза Малоука об особом «жизненном веществе» подверглась справедливой критике (Calder, 1976; Шмидт-Ниельсон, 1987). Нам также кажется, что зависимость максимальной продолжительности жизни позвоночных от относительной величины мозга имеет иную гораздо более глубокую причину, чем гипотетическое «жизненное вещество». Дело в том, что большая относительная величина мозга открывает лучшие возможности для более точной и экономной регуляции и управления системами жизнеобеспечения организма. В искусственных системах (приборах и аппаратах), чем совершеннее регуляция и управление, тем надежнее эти системы и тем продолжительней время их эксплуатации. То же самое, по-видимому, относится к живым организмам: чем лучше их управляющее устройство, что в нашем случае соответствует большей величине коэффициента энцефализации, тем надежнее живая система и тем больше продолжительность ее жизни.
Учитывая сказанное, по сопоставимой продолжительности жизни о степени организованности животных можно судить с большой осторожностью.
Эволюция максимальной продолжительности жизни животных.
Lмaх=5.39•ехр(0.0043•tg) (166) Для эволюции максимальной продолжительности жизни животных согласно данным,…Эволюция максимальной продолжительности жизни в отдельных классах позвоночных животных.
Рассмотрим вопрос о возможности использования величины максимальной продолжительности жизни для характеристики эволюционного прогресса в пределах отдельных классов. Начнем с млекопитающих. В табл.48
представлены данные о 320 видах (n=482) из 16 отрядов класса млекопитающих (Зотин,1993 а,б).
Таблица 48
Средняя масса М(кг), средняя максимальная продолжительность жизни Lmax (годы), сопоставимая максимальная продолжительность жизни 1т (годы) и время возникновения в разных отрядах млекопитающих tg (млн лет назад). N - число видов, n - число измерений.
| Таксон | M | lmax | Lm | N | n | Ч |
| Отряд Marsupialia | 4.8 | 8.6 | 3.00±0.30 | |||
| Отряд Chiroptera | 0.044 | 4.1 | 2.64±0.40 | |||
| Отряд Insectivora | 0.29 | 4.9 | 2.52±0.33 | |||
| Отряд Rodentia | 0.8 | 6.8 | 3.34±0.18 | |||
| Отряд Edentata | 5.06 | 12.2 | 4.11±0.76 | |||
| Отряд Hyracoidea | 2.3 | 10.3 | 3.63 | |||
| Отряд Cetacea | 562.1 | 17.8 | 3.58±0.65 | |||
| Отряд Lagomorpha | 1.9 | 12.2 | 4.06±0.84 | |||
| Отряд Artiodactyla | 172.3 | 21.5 | 4.35±0.12 | |||
| Отряд Carnivora | 28.6 | 18.7 | 5.23±0.20 | |||
| Отряд Sirenia | 424.0 | 30.0 | 5.08 | |||
| Отряд Tylopoda | 300.8 | 31.5 | 5.79±0.19 | |||
| Отряд Pinnipedia | 135.4 | 24.3 | 5.88±0.73 | |||
| Отряд Perissodactyla | 436.3 | 41.5 | 7.11±0.28 | |||
| Отряд Proboscidea | 3264.8 | 65.2 | 8.28±0.95 | |||
| Отряд Primates | 16.7 | 31.7 | 9.35±0.41 | |||
| Отряд Primates (без Сем. Hominidae) | 3.3 | 20.8 | 8.35±0.26 | |||
| Сем. Hominidae | 62.5 | 95.0 | 20.95±0.08 |
Как видно из табл. 48, среди млекопитающих по максимальной продолжительности жизни выделяются люди. Возможно, что это связано с
большим относительным размером мозга. Расчеты, основанные на данных, приводимых в табл.48, показывают, что эволюционный прогресс сопровождается увеличением максимальной продолжительности жизни, которую можно выразить для млекопитающих в виде:
| Lmax= 18.29 •exp(0.010•tg); | (167) |
| lm=5.46•exp(0.0053•tg). | (168) |
Формулы (167, 168) показывают, что согласно критерию, максимальной продолжительности жизни, в процессе эволюции млекопитающих совершенствование организованности происходило с той же скоростью, что и биоэнергетический прогресс и прогрессивная эволюция (143).
Таблица 49
Средняя масса М(кг), средняя максимальная продолжительность жизни Lmax (годы), сопоставимая максимальная продолжительность жизни 1т (годы) и время возникновения в разных отрядах птиц tg (млн лет назад), n - число измерений.
| Таксон | M | Lmax | lm | n | Ч |
| Отряд Galliformes | 0.928 | 7.3 | 2.40±0.17 | ||
| Отряд Pelecaniformes | 2.243 | 22.0 | 5.72 | ||
| Отряд Sphenisciformes | 18.250 | 31.5 | 5.87 | ||
| Отряд Charadriiformes | 0.174 | 16.0 | 6.79±0.37 | ||
| Отряд Falconiformes | 0.927 | 21.4 | 6.64±0.91 | ||
| Отряд Anseriformes | 3.234 | 24.3 | 6.60±0.58 | ||
| Отряд Columbiformes | 0.281 | 17.7 | 6.90 | ||
| Отряд Ciconiiformes | 1.541 | 24.3 | 6.93±0.28 | ||
| Отряд Passeriformes | 0.107 | 14.9 | 7.68±0.52 | ||
| Отряд Lariformes | 0.428 | 29.5 | 11.03±1.00 | ||
| Отряд Procellariiformes | 3.83 | 30.6 | 8.45±0.89 |
| Отряд Strigiformes | 1.208 | 34.5 | 10.12 |
С другой стороны, для класса птиц расчеты (по данным табл. 49) показывают, что максимальная продолжительность жизни и сопоставимая максимальная продолжительность жизни у этих животных практически не зависит от времени их возникновения:
Lmax=18.97•exp(-0.0029•tg) (169)
lmax=7.18•exp(0.0020·tg) (170)
В то же время для всех животных, как позвоночных, так и беспозвоночных, судя по формуле (166), увеличение степени организованности в процессе эволюции, биоэнергетический прогресс и собственно прогрессивная эволюция животных шли приблизительно с одинаковой скоростью. Эти выводы, безусловно, являются предварительными. По мере накопления и уточнения данных о максимальной продолжительности жизни и времени возникновения отдельных таксонов животных они будут изменены или уточнены. Однако принципиальная значимость для проблем прогрессивной эволюции полученных результатов очевидна.
Глава 3. Относительные размеры и энергетика мозга
Очевидно, что наиболее прямые показатели степени организованности животных должны быть связаны с совершенством нервной системы и высшей нервной деятельности. Но, к сожалению, в настоящее время нет достаточно апробированных критериев того и другого. Единственными реальными, хотя и косвенными, показателями совершенства высшей нервной деятельности животных является относительные размеры головного мозга, развитость коры больших полушарий и энергетика мозга.
Интуитивно кажется очевидным, что чем больше относительные размеры мозга, тем совершенней система управления всеми процессами в организме, тем совершенней высшая нервная деятельность животных. И действительно, к
выводу, что уровень энцефализации имеет прямое отношение к рассудочной деятельности животных и человека, приходят многие авторы (Крушинский, 1977; Крушинская, Лисицына, 1983; Jerison, Jerison, 1988). Никитенко (1966, 1969) считал, что относительные размеры мозга и коры больших полушарий являются важнейшими показателями морфо-физиологического прогресса млекопитающих. Сахер (Sacher, 1976) пошел еще дальше, считая, что lgcbr [где сbr коэффициент энцефализации (171)] является мерой содержания информации, и увеличение коэффициента энцефализации означает увеличение содержания информации в мозгу.
Относительные размеры мозга - это легко измеряемый показатель (во всяком случае, у позвоночных животных). По этому поводу накоплен большой анатомический материал не только у ныне живущих, но и вымерших животных. Ниже мы попытаемся использовать этот показатель для характеристики прогрессивной эволюции.
Кроме относительных размеров мозга для характеристики прогрессивной эволюции можно было бы использовать и данные об энергетике мозга. Но их в настоящее время немного. А жаль, так как такие материалы можно было бы использовать не только для позвоночных, но и беспозвоночных животных.
Зависимость массы мозга от массы тела.
(171) где Мbr - масса головного мозга (г); M - масса тела (г); cbr - коэффициент… Для примера на рис.39 показана зависимость массы мозга от массы тела, основанная на измерении 883 видов млекопитающих…Зависимость массы мозга от потребления кислорода
(176) Другими словами, чем больше размеры мозга, тем выше уровень потребления… устройство, чем надежней хранится и используется информация, тем больше энергии необходимо затратить на процесс…Изменение организованности животных в процессе эволюции
В настоящее время мы можем судить о величине коэффициента энцефализации только у позвоночных животных, и в этой области получены довольно… Казалось бы приведенные данные настолько демонстративны, что все ясно и по величине коэффициента энцефализации можно характеризовать эволюционный прогресс не только внутри отдельных…Часть 4. МАКРОСКОПИЧЕСКИЕ МЕХАНИЗМЫ БИОЭНЕРГЕТИЧЕСКОГО ПРОГРЕССА
С чего же начинается биоэнергетический прогресс, почему он вообще возникает, каковы предпосылки, ведущие к необходимости усиления энергетического… Прежде всего, эти предпосылки связаны с неуспехом в борьбе за существование… видами, входящими в состав биоценоза, меньше. При этом конечно появляется необходимость приспособиться к новым, часто…Глава 1. Уменьшение размеров тела
Как отмечено выше, есть несколько путей преодоления неудачной ситуации, сложившейся в биоценозе, на пути биологического процветания той или иной группы организмов. Один из них связан не с пространственной миграцией этих организмов в места с не столь напряженной конкуренцией, а с уменьшением размеров тела. Можно себе представить, что представители некого вида животные или растения, не выдерживая конкуренции в борьбе за жизнь, постепенно мельчают, так как мелкие особи имеют шанс уцелеть в результате ли потери интереса к ним основных врагов, расширения ли их кормовой базы или возможности спрятаться при возникновении опасности. Проследим, какие же изменения могут произойти при уменьшении размеров организмов.
Прежде всего, уменьшение размеров тела приводит согласно формуле (47) к усилению интенсивности потребления кислорода. Если при этом меняющиеся организмы оказываются в такой нише биоценоза, где повышенный энергетический обмен дает решающее преимущество в борьбе за жизнь, то они прекращают процесс изменений в направлении уменьшения размеров и быстро увеличивают свою численность. Если же увеличение интенсивности
стандартного обмена не дает виду заметных преимуществ (не только энергетический обмен, но и многочисленные морфологические, функциональные и поведенческие особенности организмов определяют их конкурентоспособность), то вид продолжает изменение в сторону уменьшения размеров. В конце концов, может возникнуть ситуация, показанная на рис.44. На этом рисунке приведена зависимость интенсивности потребления кислорода от массы тела у млекопитающих. Кривая, соответствующая соотношению (47), которое для млекопитающих имеет вид (Зотин, 1988):
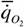 = 20.9М-0.26,
= 20.9М-0.26,
или (177)
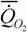 = 20.9М-0.74,
= 20.9М-0.74,
и носит название кривой Клайбера. Как видно из рис.26 она справедлива для всех млекопитающих, кроме насекомоядных [на схеме рис. 44 кривая (1)]. Расчет по данным, приводимым на рис.26, показал, что для насекомоядных аллометрическая зависимость определяется выражением:
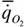 = 90,5М-0.60,
= 90,5М-0.60,
или (178)
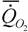 = 90.5М-0.40,
= 90.5М-0.40,
Теперь, на примере кривых, показанных на рис.44, продолжим рассмотрение гипотетического случая, когда под прессом естественного отбора происходит уменьшение размеров тела взрослых животных. По мере уменьшения массы тела в соответствии с кривой Клайбера [рис.44(2)] стандартный обмен млекопитающих возрастает. Это возрастание, начиная с массы тела порядка 20-30 г, может пойти в двух направлениях:
1) продолжить движение по кривой Клайбера [рис.44(2)],
2) вступить на более быстрый путь изменений [рис.44(1)].
В первом случае, если говорить об энергетическом обмене, ничего не изменится, так как сопоставимый стандартный обмен, т.е. коэффициент а из формулы (47), остается неизменным. Преимущества, которые при этом получают животные, связаны только с тем, что они занимают нишу, недоступную для других млекопитающих. Обладая повышенным по сравнению со всеми другими животными сходного размера (кроме птиц) стандартным обменом (рис.26) и связанными с этим возможностями, они могут найти свою нишу в биоценозе и прекратить дальнейший процесс уменьшения размеров. В смысле эволюционного прогресса уменьшение размеров тела в данном случае ничего не дает, так как критерий упорядоченности Сr, как и величина сопоставимого стандартного обмена, остаются прежними.
Во втором случае возможны два варианта.
Первый вариант. По мере уменьшения размеров организмов происходит изменение коэффициентов аллометрической зависимости (47) и стандартный обмен меняется по кривой 1, а не 2 (рис.44), т.е. возрастает значительно быстрее с уменьшением размеров тела чем по кривой Клайбера. При таком варианте животные получают значительные преимущества в величине энергетического обмена по сравнению с родительскими видами, а критерий упорядоченности у них намного больше, чем у всех остальных представителей данного класса. Для семейства землероек из отряда насекомоядных Сr=155 против Сr=104 (125) для всех остальных отрядов класса млекопитающих.
Сходные закономерности наблюдаются и среди класса птиц. Коэффициент к в аллометрической зависимости дыхания от массы тела птиц равен 0.71, в то время как в отряде калибриобразных 0.44 (126). Соответственно, в отряде калибриобразных Сr=286 против Cr=112 для всех остальных птиц (127).
Можно думать, что возникновение отряда насекомоядных в классе млекопитающих и отряда калибриобразных в классе птиц связано с изменением коэффициентов в аллометрическом соотношении (47). Это приводит к тому, что при уменьшении массы тела ниже 50-70 г стандартный обмен возрастает значительно быстрее, чем он должен был бы возрастать по кривой Клайбера, так
как аллометрическая кривая у насекомоядных и калибриобразных значительно круче, чем у остальных представителей классов, к которым относятся эти отряды [см., например, рис.44(2)]. Последнее определяет главный недостаток такого механизма прогрессивной эволюции: у крупных животных, вставших на указанный путь изменения энергетического обмена, несмотря на очень большую величину коэффициента а, а, следовательно, и критерия упорядоченности, стандартный обмен оказывается довольно низким. Расчет по формуле (178)

|
преимущество мелких насекомоядных в отношении величины стандартного обмена теряется по мере возникновения новых более крупных видов: начиная с массы в 70 г, стандартный обмен представителей этого отряда становится заметно меньше, чем у представителей других отрядов млекопитающих. Следовательно, повышенный энергетический обмен представителей нового таксона, возникшего в результате уменьшения величины коэффициента к в аллометрическом соотношении (46), ограничен областью мелких животных, т.е. имеется верхний предел размеров организмов для этого таксона. Снизу он также ограничен некоторой минимальной величиной, зависящей от возможности добычи и усвоения пищи. Дело в том, что повышенный энергетический обмен требует повышенного притока пищи. Известно, например, что мелкие землеройки из отряда насекомоядных потребляют за сутки количество пищи по массе больше, чем масса самого животного. Практически вся их жизнь состоит в непрерывном поиске пищи. Чтобы уменьшить суточную величину энергетических расходов они, подобно колибри и некоторым мелким рукокрылым, время от времени впадают в состояние оцепенения с очень низкой величиной энергетического обмена. Очевидно, что дальнейшее уменьшение
массы тела подобных животных может привести к полному прекращению активной жизнедеятельности. Для насекомоядных таким пределом является, по-видимому, масса тела меньше 2 г.
Таким образом, резкое возрастание энергетического обмена при уменьшении размеров тела в результате изменения коэффициентов в аллометрическом соотношении (46), (47) дает заметное преимущество видам, вставшим на этот путь арогенеза, но ограничивает их размеры довольно узкими рамками. Они попадают как бы в эволюционную ловушку (Зотин и др.,2000) и во всяком случае не способны вытеснить породившие их виды, имеющие массу тела выше верхнего предела ловушки.
Второй вариант. Это вариант арогенеза на начальных этапах совпадает с первым. Как и в последнем, по мере уменьшения размеров организмов происходит изменение аллометрической зависимости (47) и стандартный обмен меняется по кривой 1 [рис.44]. Это первый шаг на пути прогрессивной эволюции, позволяющий достигнуть величины сопоставимого стандартного обмена значительно большей, чем у породивших их видов. Происходит он в результате увеличения коэффициента b в соотношении (47). Второй шаг состоит в возвращении величины коэффициента b к прежнему значению при одновременном сохранении повышенной величины коэффициента а. В этом случае первоначальная кривая зависимости интенсивности дыхания от массы тела [рис.44(2)] должна сместиться вверх и все новые виды животных, независимо от их массы, будут иметь сопоставимый стандартный обмен выше, чем предковые виды. Возможным примером такого пути прогрессивной эволюции является класс птиц (рис.27).
Хорошо известно, что воробьиные птицы имеют стандартный обмен заметно выше, чем неворобьиные (Calder, 1984; Gavrilov, Dolnik, 1985; Шмидт-Ниельсен, 1987; Зотин, 1988). Расчет методом наименьших квадратов по данным, приводимым в обзоре В.М.Гаврилова и В.Р.Дольника (Gavrilov, Dolnik, 1985), показал (Зотин, 1988), что аллометрическая зависимость интенсивности потребления кислорода от массы тела у воробьиных птиц равна
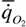 = 37.2M-0.28, (179)
= 37.2M-0.28, (179)
а у неворобьиных
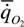 = 30.7М-0.29, (180)
= 30.7М-0.29, (180)
Соответственно, критерий упорядоченности у воробьиных птиц, согласно формуле (127), равен Сr=116.4, а у неворобьиных птиц - Сr=107.5. Так как коэффициент b y этих двух групп птиц практически одинаков, то стандартный обмен воробьиных птиц независимо от их размера всегда будет выше, чем стандартный обмен неворобьиных птиц той же массы. Это наглядно видно из рис.27, где приведены данные о стандартном обмене птиц разной массы и схематически показано на рис.44.
Некоторые авторы выделяют отряд воробьиных в отдельный подкласс, так как по численности видов, родов и семейств воробьиные превосходят всех остальных птиц вместе взятых. Это явно процветающая группа птиц наглядно демонстрирует преимущество повышения энергетического обмена и возможность на этой основе возникновения нового крупного таксона порядка отряда или даже класса.
Подводя итоги, можно сказать, что одноступенчатый механизм прогрессивной эволюции, возникающий при уменьшении массы тела животных, связанный с изменением коэффициента b в аллометрической зависимости (47), приводит к макроэволюционным изменениям животных в пределах эволюционной ловушки. Очевидно, что эволюционная ловушка может образовываться не только в процессе увеличения энергетического обмена при уменьшении размеров организмов, но и в результате чрезмерной специализации отдельных органов животных и растений, при переходе к паразитическому образу жизни и т.д.
Двухступенчатый механизм прогрессивной эволюции, первая ступень которого состоит в увеличении коэффициента b, a вторая - в стабилизации коэффициента а на высоком уровне при одновременном возвращении коэффициента b к начальному значению, приводит к макроэволюционным процессам образования крупного таксона животных или растений, неограниченных по размерам и эволюционным возможностям. Этот новый таксон обладает значительными
преимуществами в отношении энергетического обмена и способен вытеснить или, вернее, потеснить предковые формы во всех звеньях биоценоза.
Глава 2. Генотипические адаптации
Второй путь отступления видов животных и растений, терпящих поражение в борьбе за жизнь, состоит в постепенной миграции представителей этих видов на границы биоценоза, где напряженность конкуренции с более совершенными соперниками ослабевает. Вместе с тем давление отбора здесь не только не ослабевает, но значительно усиливается в результате ухудшения условий существования или появления новых соперников. Мы рассмотрим подробнее только ситуацию, когда давление отбора на границах биоценоза возрастает в результате неблагоприятных условий среды: пониженной среднегодовой температуры в северных широтах, низкого парциального давления кислорода и низкой температуры в горах на достаточно больших высотах и т.д. Способность выжить в таких новых и суровых условиях зависит от способности организмов к адаптации и последующему закреплению этих адаптации в потомстве. При этом возможны три направления изменений: два из них (аллогенез и катагенез) ведут в эволюционную ловушку, так как специализированные к плохим условиям существования организмы не имеют больших преимущества перед организмами прежних мест обитания и виды, приспособившиеся к новым условиям, не смогут завоевать утраченные позиции их предков. Другое дело когда эволюция неудачливого вида в плохих условиях существования пойдет по пути арогенеза, т.е. по пути усиления энергетического обмена. В этом случае возникающие новые виды по многим признакам (мощность, точность и скорость действий, выносливость и т.д.) окажутся на голову выше своих прежних соперников и эти новые виды способны вернуться в потерянные области прежней жизни, став основателями новых крупных таксонов.
Усиление энергетического обмена в новых условиях существования вида с термодинамической точки зрения можно охарактеризовать изменением соотношения (46). Для терпящего бедствие вида, оттесненного к границам
биоценоза, особенно важным является изменение  -функции [функции
-функции [функции
внешней диссипации индуцибильно-адаптивного процесса (56)],
характеризующей возможность популяции организмов приспособиться к новым более суровым условиям существования. Функция 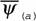 популяции
популяции
складывается из индуцибильно-адаптивных процессов, происходящих с отдельными членами популяции (57). Поэтому для того, чтобы разобраться с формированием новой популяции из уцелевших особей, мигрировавших на границу ареала, необходимо остановиться на процессе адаптации организмов к новым условиям существования. В экологической физиологии процесс адаптации организмов разбивают на три понятия: акклимация, акклиматизация и генотипическая адаптация. Все они представляют собой стационарный переходной процесс и отличаются в основном временем протекания: акклимация длится дни и недели, акклиматизация - месяцы и годы, генотипическая адаптация - несколько поколений. Хотя до сих пор не удается точнее охарактеризовать эти три понятия, они могут привести к появлению потомства с измененным генотипом, т.е. животных или растений приспособленных к новым условиям существования. Что касается прогрессивной эволюции, то это означает появление новых видов с повышенным уровнем стандартного обмена.
Рассмотрим фактический материал, имеющийся в литературе, показывающий возможность усиления стандартного обмена животных при адаптации к суровым условиям существования.
Адаптация к низким температурам среды
метаболитическая адаптация (Проссер, 1977а,б; Ивлева, 1981; Шмидт-Ниельсен, 1982). Пойкилотермные животные. В отношении пойкилотермных животных представление о… Наличие метаболитической адаптации показано также для амфибий (Владимирова, Зотин, 1994). Так, в отряде Anura наземные…Адаптация к высокогорью
обмена (Lustick, 1969; De Jong, 1976). Наоборот, понижение температуры воздуха в высокогорье должно приводить к усилению интенсивности потребления… Отделить все эти факторы друг от друга непросто, и нет ничего удивительного в… Следует учитывать, что опыты в барокамерах не являются достаточно убедительными, так как обычно животных и людей…Адаптация к физическим нагрузкам
Возрастание энергетических возможностей организмов при длительных физических нагрузках подтверждается некоторыми измерениями, проведенными при… Хорошо известно, что в результате длительных тренировок происходит возрастание… Таким образом, как и в случае адаптации животных и человека к низким температурам или горным условиям жизни, при…Глава 3. Неотения
Возможен еще один путь усиления интенсивности потребления кислорода у возникающей более прогрессивной группы животных - путь неотении, т.е. переход к активной самостоятельной жизни на ранних стадиях постэмбрионального, а иногда и поздних стадий эмбрионального развития (Зотин, Криволуцкий, 1982). Известно, что интенсивность дыхания на эмбриональных и личиночных стадиях развития животных значительно выше, чем во взрослом состоянии (Зотин, Зотина, 1993). Поэтому неотения приводит к появлению животных с более высоким уровнем стандартного обмена по сравнению с предковыми формами.
В истории сравнительной анатомии беспозвоночных на протяжении последних 150 лет было высказано немало предположений о происхождении отдельных крупных таксонов от преимагинальных стадий других групп животных. В случае насекомых, например, их происхождение от ранних стадий развития многоножек сомнений не вызывает. Этому соответствует и более низкий уровень интенсивности потребления кислорода взрослыми формами многоножек по сравнению с насекомыми: в классе Myriapoda a=0.45, Cr=5.78 (92), в то время как даже у низших насекомых отряда Colombota a=0.63, Cr=8.32 (97).
Считается, что не только насекомые и некоторые другие группы беспозвоночных возникли путем неотении, но и некоторые группы позвоночных (Воробьева, 1992; Иорданский, 1994) и многие группы растений (de Beer, 1958; Тахтаджан, 1970; Капанадзе, Капанадзе, 1995).
Явление неотении в случае измельчания вида или перехода его в неблагоприятные условия существования связано с трудностями борьбы за существование взрослых форм животных. Дальнейшая эволюционная судьба
неотенического вида, как и в случае измельчания, зависит от способности животных изменить коэффициенты в аллометрическом соотношении (46). В этом случае возможны три пути изменения энергетического обмена у возникающих групп животных (рис.38). Два из них, когда 0.75<k<1.00 и когда k=0.75 (если при этом возрастает коэффициент а), ведут по пути биоэнергетического прогресса, один, когда k<0.75, ведет в эволюционную ловушку. Неотенические виды имеют преимущество в энергетическом обмене при малых массах и недостаток - при больших по сравнению с предковыми формами.
В настоящее время существуют и процветают животные, которые не перешли полностью на путь неотении, но уже способны размножаться на личиночных стадиях развития. Другими словами, они могут существовать и давать потомство, не переходя во взрослое состояние. Стоит им потерять взрослые формы, и они превратятся в новый вид, взрослыми стадиями которого будут личинки предков.
Это, прежде всего, относится к семейству Ambystomidae из отряда хвостатых амфибий. Личинки этих амфибий - аксолотли - способны, как и взрослые формы - амбистомы - к размножению. Так как на личиночных стадиях развития интенсивность потребления кислорода выше, чем у взрослых форм, то к моменту половозрелости интенсивность дыхания аксолотлей выше, чем амбистом (Владимирова и др., 1993). Если бы представители рода Ambystoma полностью перешли на личиночный тип жизни, то образовавшийся новый вид обладал бы большей величиной коэффициента а и критерия упорядоченности.
Подтверждение этому выводу находим в работах Владимировой, Сербиновой (1992), Владимировой и др.(1993), в которых показано, что интенсивность потребления кислорода хвостатыми амфибиями в 2 раза ниже, чем бесхвостых (рис.24). В то же время, исследованный авторами вид Ambystoma mexicanum по уровню стандартного обмена занимают двоякое положение: у половозрелых амбистом, как показывает расчет по данным Владимировой и др.(1993), а = 0.22, в то время как у половозрелых аксолотлей а = 0.49 мВт (табл.31). Другими словами, амбистомы стоят в системе хвостатых
амфибий на первом месте, а половозрелые неотенические личинки - аксолотли - на последнем и не отличаются по стандартному обмену от некоторых бесхвостых амфибий (табл.35). То же свойственно и некоторым другим видам хвостатых амфибий, имеющих способных к размножению неотенических личинок (Bentley, Shield, 1973; Shield, Bentley, 1973; Ultsch, 1974; Bentley, 1975; Withers, Hillman, 1981). Интенсивность дыхания половозрелых личинок этих амфибий, как и в случае аксолотля, близка к интенсивности дыхания бесхвостых амфибий. Таким образом, в этих группах животных формируется новый таксон класса амфибий, который по уровню энергетического обмена приближается к отряду бесхвостых амфибий.
Интересные результаты были получены при изучении интенсивности потребления кислорода при метаморфозе аксолотлей в амбистомы (Владимирова и др., 1993). Метаморфоз аксолотлей вызывали на разных стадиях роста добавлением в воду тиреоидина в концентрации 1 г/мл, как рекомендует Воронцова и др. (1952). Потребление кислорода измеряли монометрическим методом Варбурга при 20°С в специально изготовленных сосудах цилиндрической формы. Была исследована интенсивность потребления кислорода аксолотлями от стадии вылупления (масса тела 0.01 г) до максимальной массы тела в 130г.
Как видно из рис.47 интенсивность дыхания аксолотлей непрерывно уменьшается во время развития. Можно выделить два периода изменения интенсивности дыхания: до 2 г и после, - когда уже возможен метаморфоз аксолотлей в амбистом. Поэтому, сравнивать дыхание аксолотлей и амбистом лучше во втором возрастном периоде. Половозрелые аксолотли имели массу тела в среднем равную 77 г и интенсивность потребления кислорода 0.202 мВт/г. Половозрелые амбистомы имели массу тела в среднем 34 г и интенсивность потребления кислорода 0.112 мВт/г
Аллометрическая зависимость скорости потребления кислорода от массы тела во втором возрастном периоде у аксолотлей имеет вид
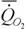 = 0.139·М0.87 (181)
= 0.139·М0.87 (181)
У амбистом на всех стадиях роста от 3 до 50 г интенсивность потребления кислорода была ниже, чем у растущих аксолотлей сходной массы (рис.47). Аллометрическая зависимость скорости дыхания от массы тела у амбистом имела вид:
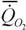 = 0.103М 0.49 (182)
= 0.103М 0.49 (182)
Из сопоставления этих формул видно, что коэффициент а у аксолотлей на 30% выше, чем у амбистом. Оказалось, что это справедливо и для межвидовой аллометрии, так как стандартный обмен аксолотлей разных видов примерно на 30% выше, чем у амбистом. Можно думать, поэтому, что увеличение коэффициента а и критерия упорядоченности у аксолотлей произошло за счет уменьшения коэффициента k [сравните аллометрические соотношения (181) и (182)]. А это означает, что только в том случае, когда коэффициент k для растущих личинок выше, чем у взрослых, неотения может привести к появлению видов с более высоким уровнем биоэнергетики. Возможно, что увеличение коэффициента а у насекомоядных млекопитающих (125) и колибриобразных птиц (127) произошло тем же путем, что и у аксолотлей. Не исключено, поэтому, что в основе образования указанных отрядов животных лежала неотения, связанная с переходом к взрослому существованию на ранних стадиях роста предков.
Глава 4. Митохондриальные механизмы прогрессивной эволюции
Подводя итоги рассмотрению различных механизмов изменения энергетического обмена в процессе эволюции, следует отметить, во-первых, что все они могут приводить не только к повышению стандартного обмена (арогенез) или сохранению его прежней величины (аллогенез), но и к понижению (катагенез), во-вторых, во всех случаях в основе
макроэволюционных механизмов лежат микроэволюционные изменения. Это касается и арогенеза, т.е. механизмов биоэнергетического прогресса.
Специфичность механизмов биоэнергетического прогресса и прогрессивной эволюции состоит в том, что в обоих случаях речь идет о совершенствовании энергетики клеток и, в первую очередь, митохондриального аппарата. Как показано выше, специфичность механизмов биоэнергетического прогресса определяются тем, что виды, проигрывающие битву за существование, вынуждены или уменьшать свои размеры, или увеличить физические нагрузки (например, в результате быстрых и длительных перемещений), или мигрировать в холодные географические районы и горы, или вступать на путь неотении. Если в этих условиях у них вырабатываются приспособления, повышающие стандартный обмен, и он закрепляется генетически, они переходят на новый уровень энергетического обмена, который позволяет им (или порожденным ими видам) завоевать утерянные позиции и лечь в основу макроэволюционного процесса. Кажется очевидным, что основным приспособлением, повышающим стандартный обмен, может служить увеличение числа митохондрий в клетках и совершенствование их функционирования.
Доказательство того, что указанные выше макроскопические механизмы прогрессивной эволюции сопровождаются увеличением концентрации митохондрий в клетках и тканях животных приведены в нескольких монографиях (Меерсон, 1973, 1981; Зотин, 1988; Озернюк, 1992). Меерсон (1973, 1974), в частности, считал, что процесс адаптации к холоду, гипоксии и физическим нагрузкам сопровождается увеличением содержания митохондрий в единице массы клеток и тканей (рис.48). Согласно схеме Меерсона под действием указанных факторов происходит снижение содержания АТФ в клетках, что приводит к нарушению жизнедеятельности организма, активации генетического аппарата, усилению биогенеза митохондрий и восстановлению нарушенного уровня содержания АТФ.
Температурная адаптация. Имеется некоторое число исследований, подтверждающих идею Меерсона относительно роли биогенеза митохондрий в
процессе адаптации животных к холоду. Так, активность многих митохондриальных ферментов в гомогенатах печени, мышц и других органах крыс, адаптированных к холоду, повышена (Меерсон, 1973, 1974). Показано также, что активность цитохромоксидазы печени, мозга, мышц и жабр золотых рыбок, акклимированных к 10°С, на 20-27% выше, чем у рыбок, акклимированных к 30°С (Caldwell, 1969), а в мышцах рыб, акклимированных к 8°С, на 30% выше, чем у акклимированных к 22°С (Jankowsky, Korn, 1965).
Довольно много данных об изменении структуры и содержания митохондрий при адаптации рыб к пониженным температурам среды приведено в монографии Озернюка (1992). Так, было установлено, что при акклимации золотых рыбок к температуре 5°С активность цитохромоксидазы в скелетных мышцах на 66% выше, чем при акклимации к 25°С. То же самое относится к такому митохондриальному ферменту, как сукцинатдегидрогеназа. Показано, что содержание цитохрома с в скелетных мышцах солнечной рыбы Lepomis ceantllus при акклимации к 5°С равнялась 1.51 нмоль/г, при 15°С - 1.17 и при 25°С - 0.98 (Sidell, 1977), т.е. при адаптации к 5°С содержание цитохрома с в мышцах повышается на 29% по сравнению с рыбами, адаптированными к 15°С, и на 54% - по сравнению с рыбами, адаптированными к 25°С.
Подобные данные получены о содержании цитохромов аа3, с1 с и b в красных и белых мышцах рыбок Tilapia mossambica, акклимированных в течение 20 дней к температуре 20, 26 и 32°С (рис.49). Оптимальной для тиляпий является температура в 26°С. Как видно из рис.49, понижение температуры акклимации до 20°С приводит к резкому увеличению содержания цитохромов в скелетных мышцах теляпий - особенно в красных мышцах. Так как цитохромы дыхательной цепи входят в состав внутренней мембраны митохондрий, полученные результаты свидетельствуют о том, что в процессе холодовой адаптации усиливается биогенез митохондрий (Озернюк, 1992). Об этом свидетельствуют также данные об увеличении относительной массы митохондрий в скелетных мышцах рыб при адаптации к холоду. Так, у карпов, акклимированных к 2°С, митохондрии занимают 25% объема мышечных волокон красных мышц, в то время как у акклимированных к 28°С только 14%
(Johnston, Maitland, 1980). В красных мышцах полосатого окуня при акклимации к 25°С относительная площадь митохондрий составляет 29%, а при акклимации к 5°С - 45% (Egginton, Sidell, 1986).
Таблица 58 Относительная площадь митохондрий в скелетных мышцах рыб (Dunn, 1988).
| Виды | Красные мышцы | Белые мышцы |
| Антарктические рыбы | ||
| Chaenocephalus aceratus | _ | |
| Notothenia gibberifrons | _ | |
| Notothenia rossi | 1.9 | |
| Pleurogramma antarcticum | 6.3 | |
| Рыбы с низкой двигательной активностью | ||
| Anguilla ro strata | 1.0 | |
| Carassius carassius | 1.6 | |
| Ciarias mossambicus | 2.6 | |
| Conger conger | 2.7 | |
| Pleuronectes platessa | 2.0 | |
| Protopterus aethiopicus | 0.2 | |
| Рыбы с высокой двигательной активностью | ||
| Engraulis encrasicolus | _ | |
| Katsuwonus pelamis | 2.3 | |
| Pollachius virens | 5.1 | |
| Salvelinus fontinalis | 9.3 | |
| Scomber scomder | _ |
Возможность генетического закрепления повышенного содержания митохондрий при холодовой адаптации животных показано на примере антарктических рыб, относительный объем митохондрий которых, судя по некоторым наблюдениям (табл.58), больше, чем у теплолюбивых рыб (Озернюк. 1992).
Таким образом, адаптация к низким температурам может завершиться закреплением повышенного содержания митохондриальных мембран или митохондриальных белков, и, следовательно, увеличенного стандартного обмена у животных.
Высокогорье. Вторым фактором среды согласно схеме Меерсона (рис.48), вызывающим увеличение содержания митохондрий в клетках и тканях животных, является гипоксия. Высоко в горах действует также понижение температуры, повышенный уровень радиации и т.д. (Зотин, 1988). Несмотря на сохранение или даже снижение уровня потребления кислорода в этих условиях, общая мощность системы усвоения кислорода на клеточном уровне возрастает (Барбашова, 1970; Меерсон, 1973, 1981).
По мнению Меерсона (1981) это связано, в частности, с увеличением содержания митохондрий в клетках и тканях.
Физические нагрузки. Согласно схеме Меерсона (рис.48) третьей причиной увеличения концентрации митохондрий в клетках и тканях животных могут служить повышенные физические нагрузки. Имеются и прямые доказательства того, что при адаптации к большим физическим нагрузкам происходит увеличение содержания митохондрий в мышцах (Меерсон, 1973; Зотин, 1988). В частности, показано, что в скелетных мышцах белых крыс, которых в течение 8 недель заставляли плавать 2 раза в день, содержание митохондриального белка возросло на 19% (Рогозкин и др., 1964), а тренировка плаваньем в течение 160 часов привела к увеличению концентрации митохондрий в миокарде крыс на 52% (Arcos et al., 1968). Ежедневные тренировки в беге на тредбане с увеличивающейся нагрузкой на протяжении 12 недель привела к увеличению концентрации митохондриального белка в мышцах крыс на 60% и активности ферментов дыхательной цепи и цитохрома С в 2 раза (Holloszy, 1967).
Электронно-микроскопическое изучение содержания митохондрий в икроножной мышце белых крыс после 10 недель тренировки в беге показало, что количество митохондрий возросло на 50-100% по сравнению с контролем (Gollnik, King, 1969).
У рыб с высокой двигательной активностью относительная площадь митохондрий больше, чем у рыб с низкой двигательной активностью (табл.58). Было показано также, что содержание цитохромов в скелетных мышцах рыб коррелирует со скоростью плаванья: у малоподвижных рыб концентрация цитохромов заметно ниже, чем у быстро плавающих (рис.50).
* * *
Подводя итоги рассмотрения возможных механизмов биоэнергетического прогресса и прогрессивной эволюции, мы можем опереться на следующую схему. В результате неуспеха в борьбе за существование некоторые виды животных начинают уменьшаться в размерах, переходить к размножению на личиночных стадиях развития, увеличивать способность к повышенным физическим нагрузкам или мигрируют в суровые условия существования (холодные районы Земли, в горы и т.д.). В некоторых случаях это приводит к увеличению стандартного обмена за счет увеличения концентрации митохондриальных ферментов и усиления их функционирования в клетках и тканях. Если в результате этого процесса возникают новые виды с повышенным энергетическим обменом, они вступают на путь прогрессивной эволюции и могут дать начало макроэволюционному процессу, который приведет к появлению группы видов, обладающих преимуществом в борьбе за жизнь по многим показателям. Они начнут теснить бывших врагов-победителей в те же условия, в которых пребывали сами, и, тем самым, создадут условия для возникновения нового более прогрессивного макроэволюционного процесса.
Часть 5. БИОЭНЕРГЕТИЧЕСКИЙ ПРОГРЕСС И ТЕПЛОВЫЕ БАРЬЕРЫ
Очевидно, что повышение температуры тела гомойотермных животных связано с усилением энергетического обмена. Для того, чтобы доказать это, в табл.59… Если показать зависимость температуры тела млекопитающих от сопоставимого… t0 = 33.3·exρ(0.0037·a). (183)Глава 1. Первый тепловой барьер и возникновение терморегуляции
Данная глава посвящена причинам и условиям возникновения терморегуляции в процессе биологической эволюции. Но речь идет не о поведенческой терморегуляции или пассивной терморегуляции, связанной с увеличением размеров тела, изменением покровов тела и т.д. (так называемой, эктотермии), а о химической и физической терморегуляции, свойственной гомойотермным
животным (эндотермия). Под термином терморегуляция мы будем понимать только эндотермию.
Чтобы разобраться с вопросом, как терморегуляция связана с биоэнергетическим прогрессом, напомним кратко о проблеме эволюционного прогресса вообще. Согласно Северцову (1934.1939), в основе морфо-физиологического прогресса лежит повышение общей энергии жизнедеятельности организмов, т.е. как, показал Дольник (1968) и мы (часть 2 и 4), с усилением стандартного обмена животных, измеряемого по скорости потребления кислорода в состоянии покоя при 20°С у пойкилотермных и в термонейтральной зоне у гомойотермных животных. Стандартный обмен организмов связан с размерами тела животных аллометрическими соотношениями (46) и (47), поэтому, при сравнении энергетического уровня животных разных видов, имеющих разную массу, используют коэффициент а, который соответствует величине потребления кислорода животными, если бы они имели одинаковую массу равную 1 г.
В.Р.Дольник (1968) один из первых сопоставил коэффициент а у животных разных классов и типов и показал, что по мере усложнения строения и организации животных их стандартный обмен значительно возрастает (табл.1). Нами была продолжена работа по сбору данных о стандартном обмене животных, которая отражена на рис. 13-27 и табл.41. На рис.28 было показано, что в процессе прогрессивной эволюции стандартный обмен (коэффициент а) возрастает от простейших до млекопитающих и птиц в сотни раз. Это показывает, в соответствии с утверждением В.Р.Дольника, что имеет место биоэнергетическая направленность эволюционного прогресса организмов.
Как следует из термодинамики неравновесных процессов, в общем случае, система любой степени сложности в процессе изменений движется к состоянию равновесия. В то же время, в термодинамике неравновесных процессов существует, как указывалось, принцип наискорейшего спуска, согласно которому движение открытой системы к равновесию происходит самым быстрым путем. Отсюда вытекает, что движение сложной системы к равновесию или стационарному состоянию должно сопровождаться появлением подсистем,
которые двигаются в противоположном направлении и это движение сопровождается увеличением диссипативных функций подсистемы. В случае биосферы это сопровождается появлением групп организмов с повышенным энергетическим обменом. Появление таких подсистем (таких организмов) ускоряет движение системы в целом (биосферы) к состоянию равновесия.
Эта термодинамическая закономерность (принцип наискорейшего спуска) заставляет некоторые группы организмов двигаться все дальше по пути биоэнергетического прогресса, несмотря на все препятствия, которые встречаются на этом пути. Появление терморегуляции у животных в этом случае является необходимым следствием этой закономерности, так как только таким путем, после преодоления тепловых барьеров, возможно дальнейшее движение в сторону ускоренного усиления потребляемой энергии живыми системами.
Как отмечалось выше, стандартный обмен недостаточен для характеристики энергетического обмена животных, так как он характеризует энергетику в состоянии покоя, а животные значительную часть своей жизни проводят в активном состоянии. Для примера на рис.52 приведены данные об усилении интенсивности потребления кислорода при изменении скорости плаванья солнечной рыбы Lepomis gibbosus в гидродинамической трубе (Brett, Sutherland, 1965). Как видно из этого рисунка величина активного обмена может в 5-6 раз превышать стандартный обмен. Однако сравнивать между собой активный обмен разных животных довольно трудно, так как он сильно зависит от способов перемещения или совершения работы. Лучше в этом отношении обстоит дело с максимальным обменом, который соответствует интенсивности потребления кислорода животными и человеком в условиях максимальной физической нагрузки. Было показано, что максимальный обмен животных прямо пропорционален величине стандартного обмена (Шмидт-Ниельсен, 1987) и превышает последний у всех животных (рис.12) в 9.6 раза (Донцова, Зотин, 1980; Зотин, 1988).
Нас здесь интересует не вопрос о том, насколько точна эта цифра, а сам факт значительного увеличения энергетического метаболизма при активном и, тем более, максимальном обмене. То и другое является чрезвычайно важным
обстоятельством в жизни многих животных, без которых им трудно выжить в борьбе за существование. Но увеличение энергетического обмена при активном поведении животного неизбежно приводит к увеличению температуры тела. Например, температура тела во время свободного полета бражника или шмеля достигает 40°С при 16-33°С среды (Heinrich, 1971, 1975), а матки шмеля - до 45°С (Heinrich, 1975). Учитывая, что сопоставимый стандартный обмен пчел равен а=4.74 мВт/г (Алексеева, Зотин, 1996), мы можем определить эту цифру в качестве нижней границы энергетического обмена, для перехода которой необходимо появление способности к терморегуляции иначе при активном обмене образуется слишком много тепла внутри организма.
Верхнюю границу стандартного обмена, выше которой существование животных без возникновения терморегуляции затруднительно, мы можем определить исходя из данных о стандартном обмене однопроходных. Ехидны имеют слабую систему терморегуляции одну из самых несовершенных среди млекопитающих (Владимирова, Зотин, 1989), при этом сопоставимый стандартный обмен семейства Tachyglossidae равен а = 8.3 мВт (Зотин и др., 1990). Отсюда следует, что выше этой величины сопоставимого стандартного обмена температура тела животного, не способного к терморегуляции, при активном и тем более максимальном обмене может достигнуть такой величины, что оно погибнет или будет повреждено .Исходя из сказанного, мы и назвали сопоставимый стандартный обмен от 5 до 8 мВт/г первым тепловым барьером (Зотин и др., 19986).
Другой способ приблизительного определения нижней границы первого теплового барьера заключается в использовании данных о, так называемом, метаболитическом диапазоне (D) (Шмидт-Ниельсен, 1987), который соответствует превышению максимального обмена над стандартным. Как говорилось, для всех животных он приблизительно одинаков и равен D=9.6 (Зотин, 1988). Границу первого теплового барьера можно определить, деля наивысшую величину стандартного обмена в отряде воробьиных птиц на величину метаболитического диапазона, считая, что воробьиные достигли максимально возможной температуры тела и дальнейшее увеличение
стандартного обмена у гомойотермных животных нежелательно, как и максимального обмена пойкилотермных животных. Мы, сравнив кривую биоэнрегетического прогресса и кривую максимального обмена (рис.53) пришли к выводу, сходному с приведенным выше: примерно с величины коэффициента а равной 5-6 мВт/г величина максимального обмена становится больше 75 мВт/г, т.е. достигает уровня второго теплового барьера, когда температура тела поднимается до летальных значений (рис.51).
Данные о сопоставимом стандартном обмене [коэффициент а из аллометрического соотношения (46), (47)] для некоторых семейств воробьинообразных птиц, взятые из статьи Зотина (1979), приведены в табл.40. Если рассчитать по этим данным величину a/D, то оказывается, что граница первого теплового барьера лежит в районе 4-5 мВт/г, т.е. в той же области, что и полученная по данным о стандартном обмене пчел. Больше того, если учитывать верхнюю границу, полученную по данным о стандартном обмене низших млекопитающих, равную 8 мВт/г, можно думать, что у птиц еще есть некоторая возможность наращивать величину стандартного обмена в процессе эволюции.
Таким образом, для пойкилотермных животных существует некая зона стандартного обмена, равная приблизительно 5-8 мВт/г, выше которой животные нормально существовать не могут. Этот барьер пойкилотермные должны преодолеть в процессе биоэнергетического прогресса. Преодолеть его можно только в результате появления способности к терморегуляции, т.е. превращения пойкилотермных животных в гомойотермных.
На первый взгляд это несколько не совпадает с существующими представлениями об эволюции процесса терморегуляции (Слоним, 1986), но на самом деле принципиальных различий между этими двумя подходами нет, так как прогрессивная эволюция всегда поддерживается естественным отбором.
Способны ли крокодилы и динозавры к терморегуляции?
терморегуляции у вымерших млекопитающих и птиц, позволяет определить, способны ли крупные рептилии, например, крокодилы к терморегуляции и следует… Начнем с того, можем ли мы определить коэффициент а из аллометрического… (185)Глава 2. Второй тепловой барьер и возникновение цивилизаций.
Как отмечалось, в результате биоэнергетического прогресса и увеличения стандартного обмена температура тела гомойотермных животных достигает такой величины, что дальнейшее увеличение стандартного обмена становится невозможным. Биоэнергетический прогресс достигает нового, второго теплового барьера и должен был бы остановиться.
То, что увеличение сопоставимого стандартного обмена [коэффициента а из формулы (46)] коррелирует с увеличением температуры тела (коэффициент корреляции, как говорилось, равен 0.78) следует из данных, приводимых в табл.59 и на рис.51. Поэтому дальнейшее увеличение стандартного обмена, которое является неизбежным следствием принципа наискорейшего спуска термодинамики неравновесных процессов, выше определенной величины становится невозможным.
Природа, однако, нашла такой путь эволюции животных, который позволяет преодолеть и второй тепловой барьер - это путь возникновения цивилизаций. Речь идет о совершенствовании в процессе прогрессивной эволюции системы регуляции и управления в живых организмах, которая завершилась в человеке появлением сознания. Это совершенствование связано и даже базируется на биоэнергетическом прогрессе, так как согласно термодинамике информационных процессов в одинаковых по строению устройствах более совершенная система получения, хранения и использования информации, более точная работа регулирующих и управляющих систем требует больших затрат энергии (Поплавский, 1981). Отсюда следует, что возможность совершенствования информационных систем животных зависит от возможностей их энергетики: только у организмов, достигших высокого уровня энергетического обмена возможно появление совершенной системы регуляции и управления и, тем более, сознания.
Чем же сознание может помочь преодолению второго теплового барьера и осуществлению дальнейшего движения по пути биоэнергетического прогресса? Прежде всего тем, что используются источники энергии не только внутри организма, но и вне его. Это смог осуществить в больших масштабах пока только человек, который научился употреблять для своих нужд не только энергию, заключенную в пище, но и в различных горючих материалах, энергию рек, ветра и солнца, а в последние десятилетия и атомную энергию. Человечество на протяжении тысячелетий своей истории постепенно увеличивало количество используемой энергии, приходящейся на каждого индивидуума (табл.60).
Таблица 60
Потребление и производство энергии в процессе эволюции человечества (Zotin, Lamprecht, 1996)*
| Уровень цивилизации | Историческое время | Потребление энергии на одного человека | |
| годы | Ккал/день | а (мВт) | |
| Примитивный человек | -2000000 | 22.5 | |
| Использование огня | -750000 | 56.2 | |
| Доместификация животных | -6000 | 134.8 | |
| Индустриальная революция | +1800 | ||
| США (настоящее время) | +1990 | ||
| Весь мир (наст, время) | +1993 |
* По данным: Adams, 1975; Кнорре, 1978; Eppler, 1989
Вначале этот процесс происходил относительно медленно, а затем со все большим ускорением. Так, расчеты показывают (табл.60), что первобытный человек получал с пищей не более 2000 Кал/сутки, при использовании огня костров потребление энергии увеличилось до 5000 Кал/сутки, 6-7 тыс. лет тому назад - до 12000, в XVII-XIX веках (Англия) - до 77000 и в современное время
(США) - 230000 Кал/сутки. Особенно ускорился рост мирового производства энергии в XVIII-XX веках. По данным Роуза и Кларка (1963) мировое потребление энергии в 1860 году составляло примерно 0.017 ·1021 , в 1946 году - 0.093 ·1021 , а в 1975 должно было составлять 0.34 · 1021 дж.
Чтобы более наглядно показать преемственность энергетического прогресса животных, протекающего на протяжении миллионов лет, и человеческой цивилизации, происходящего в последние несколько тысяч лет, следует сравнить кривую биоэнергетического прогресса и кривую увеличения энергообеспеченности человечества. Напомним еще раз, что для сравнения энергетического обмена животных разных классов и типов животных используют коэффициент а из формулы (46). Поэтому, чтобы сравнить рост энергетических возможностей человечества с биоэнергетикой животных, приходится прибегать к сходной величине. Принимая, что в среднем масса человека равна 65000 г, а коэффициент k =0.75, формулу для расчета а в разные периоды развития человеческой цивилизации можно записать в виде:
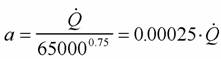 (189)
(189)
где 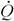 - потребление энергии в мВт/г. С помощью этой формулы и данных о энергопотреблении людей в разные периоды человеческой цивилизации (Табл.60) можно получить кривую энергетического прогресса человечества и сопоставить ее с кривой биоэнергетического прогресса животных. Как видно из рис.53(А), где приведено это сопоставление, кривая роста энергообеспечения людей как бы продолжает кривую увеличения энергетического обмена животных.
- потребление энергии в мВт/г. С помощью этой формулы и данных о энергопотреблении людей в разные периоды человеческой цивилизации (Табл.60) можно получить кривую энергетического прогресса человечества и сопоставить ее с кривой биоэнергетического прогресса животных. Как видно из рис.53(А), где приведено это сопоставление, кривая роста энергообеспечения людей как бы продолжает кривую увеличения энергетического обмена животных.
Это не может не вызвать удивления. Природа создала условия для постепенного усиления биоэнергетики отдельных групп животных и, когда возникло препятствие, связанное с возрастанием температуры тела, нашла путь обхода этого препятствия в виде появления разумной деятельности одного из видов приматов. Этот обходный путь начался задолго до приближения
животных в процессе эволюции ко второму тепловому барьеру и не только в отряде приматов.
Как уже отмечалось закономерность, которая заставляет организмы двигаться все дальше по пути биоэнергетического прогресса, несмотря на все препятствия, которые встречаются на этом пути, опирается на принцип наискорейшего спуска термодинамики неравновесных процессов. Следовательно, появление цивилизаций является необходимым следствием термодинамической закономерности, так как только таким путем, после преодоления второго теплового барьера, возможно дальнейшее движение в сторону увеличения потребляемения энергии живыми системами. Возникновение цивилизации на Земле является неизбежным результатом этой термодинамической закономерности и вытекающего из нее биоэнергетического прогресса. Отсюда следует также, что если бы не появился человек, на его месте мог оказаться некоторый другой вид животных, способный создать цивилизацию в указанном выше смысле. Что такое возможно следует из данных о возрастании массы мозга в процессе прогрессивной эволюции в разных классах и типах животных.
Увеличение относительных размеров мозга
Трудно, однако, использовать для целей сравнения относительные размеры мозга, если понимать под этим понятием отношение массы или объема мозга к… коэффициента энцефализации. Он соответствует массе мозга, если бы животное… (190);Виртуальные цивилизации.
Представители этой группы динозавров показаны на рис.55. Коэффициент энцефализации этих вымерших рептилий в 5.8 раз больше чем у современных… Интересно отметить, что наибольший коэффициент энцефализации внутри быстро… Например, среди человекообразных обезьян, занимающих одно из самых заметных мест по величине коэффициента…Социальная деятельность животных
Следует отметить, что насекомые обладают наружным скелетом, и это в значительной степени ограничивает возможности увеличения их размеров: при… иметь порядок 1000-2000 г, а относительная - Cbr=2 - 3. Эти цифры недостижимы… Речь идет о создании сообществ насекомых в виде термитников, муравейников, пчелиных гнезд и т.д. Эти сообщества…Глава 3. Третий тепловой барьер и выход человечества в космическое пространство
Многие авторы, изучающие проблемы, связанные с эволюцией биосферы, рассматривают появление человеческой цивилизации как следствие биологической эволюции жизни на Земле (Вернадский, 1926, 1940, 1967;
Верзилин и др., 1976; Вассоевич, Иванов, 1977; Камшилов, 1979; Шипунов, 1980; Водопьянов, 1981; Соколов, 1986; Будыко, 1984; Сидоренко, 984; Моисеев и др., 1985; Колчинский, 1990).
Как следует из работ Вассовича и Иванова (1983), Шиловой (1983) и Колчинского (1990) термином биосфера [введенным Зюссом (Süess, 1875)] обозначают совокупность живых организмов, обитающих на Земле, вместе с образуемыми этими организмами в прошлом и настоящем геологическими формациями.
В.И.Вернадский (1926, 1940, 1967) называл биосферой часть атмосферы, гидросферы и литосферы, строение и состав которой связаны с жизнедеятельностью организмов. Естественно, поэтому, что по мере органической эволюции происходило изменение химического состава этих слоев Земли. В свою очередь, изменение геохимической обстановки в целом и в отдельных регионах значительно влияло на процесс эволюции организмов. Несмотря на это, ведущими в процессе эволюции биосферы, по мнению большинства авторов, начиная с Ламарка, были процессы, связанные с изменениями живых систем: возникновение огромных захоронений углеродистых остатков, существенные изменения состава атмосферы и гидросферы, изменение соотношения химических элементов и формирование некоторых геологических образований в поверхности Земли, - все это результат деятельности и миграции различных видов организмов. Таким образом, органический мир детерминирует основные параметры биосферы и играет ведущую роль в ее эволюции (Колчинский, 1990).
Переломными этапами эволюции биосферы были возникновение жизни в гидросфере, переход от восстановительной атмосферы к окислительной и появление человечества. Все три этапа обусловлены эволюцией жизни на Земле. Важнейшими событиями для истории биосферы были такие события как возникновение основных способов питания (хемотрофного, автотрофного, гетеротрофного), появление организмов, обладающих скелетом, и способных к активным перемещениям, переход организмов от водного образа жизни к наземному, цефализация и связанный с этим разумный способ поведения животных и др. Последнее в конечном итоге привело к появлению человека и постепенному преобразованию биосферы в ноосферу [термин, предложенный
Леруа (Le Roy, 1927)], которая, согласно Вернадскому (1926) и другим, приводит к резкому изменению геохимической истории Земли в результате производственной деятельности человека.
Ноосфера как следствие принципа наискорейшего спуска
Мы уже рассмотрели эти вопросы в предыдущей главе и пришли к выводу, что то и другое возникло бы на Земле независимо от того, появился ли бы здесь… Почему же происходит этот процесс - процесс непрерывного увеличения энергетики… Биологический смысл биоэнергетической направленности эволюционного прогресса состоит в том, что более высокий уровень…Принцип наискорейшего спуска, как основа эволюции Вселенной
По сообщению Шкловского (1987) Н.С.Кардашев еще в 1964 году по уровню достигнутого технического развития выделил три возможных типа цивилизаций: I. Технологический уровень близок к тому, который уже достигнут на Земле.… П. Цивилизация овладела энергией, излучаемой звездой. Ежесекундное потребление энергии около 1033 эрг.Третий тепловой барьер
Следствием перспективы возникновения третьего теплового барьера является необходимость выхода человечества в космическое пространство и организации… составляло около 6·1019 эрг/с, причем в течение последних 200 лет каждые… Конечно, значительное увеличение производства энергии сопровождается и другими последствиями, неблагоприятными для…ЗАКЛЮЧЕНИЕ
Еще одной особенностью науки является то, что большей частью установленные в ней истины, вошедшие в основной фонд науки, имеют длительную историю.… иного автора, так как конечная истина базируется на ряде неясных, а порой и… Так, термодинамический подход к проблемам биологии начат еще в работах Больцмана и продолжается до наших дней. Даже…ЛИТЕРАТУРА
Аболмасова Г.И. Скорость обмена у некоторых видов беспозвоночных из
Средиземного моря // Биология моря / Ред.В.Е.Заика и др. Киев:
Наукова Думка. Киев: Наукова Думка. 1878. Вып.46. С.25-29.
Аболмасова Г.И. Скорость энергетического обмена и пищевые потребности
кальмара Sthenoteuthis oualaniensis из Индийского океана в
экспериментальных условиях // Экология моря / Ред. В.Е.Заика и др.
Киев: Наукова Думка. 1985. №19. С. 104-110.
Аболмасова Г.И., Белокопытин Ю.С. Изучение энергетического обмена у
эрипелагического кальмара Sthenoteuthis pteropus St. при разных
скоростях плаванья в экспериментальных условиях // Экология моря
/ Ред. В.Е.Заика и др. Киев: Наукова Думка. 1987. №27. С.73-78.
Аболмасова Г.И., Муравская З.А. Потребление кислорода и экскреция азота
у кальмара Sthenoteuthis pteropus St. // Экология моря / Ред. В.Е.Заика
и др. Киев: Наукова Думка. 1985. №21. С.57-60.
Аболмасова Г.И., Столбов А.Я. Изучение скорости энергетического обмена у
крылорукого кальмара (Sthenoteuthis pteropus St.) при низких
температурах с учетом гидростатического давления // Экология моря
/ Ред. В.Е.Заика и др. Киев: Наукова Думка. 1991. №37. С.73-76.
Аболмасова Г.И., Щепкина A.M., Юнева Т.В. Энергетические особенности
кальмаров // Биоэнергетика гидробионтов / Ред.Г.Е.Шульман,
Г.А.Финенко. Киев: Наукова Думка. 1990. С.83-101.
Айдаралиева A.A., Исабаева В.А., Слоним А.Д. Реакции организма на
гипоксию // Экологическая физиология животных / Ред. А.Д.Слоним
и др. Л.: Наука. 1982. Ч.З. С.308-323.
Акимушкин И.И. Класс Головоногих (Cephalopoda) // Жизнь животных. Т.2.
Беспозвоночные / Ред. Л.А.Зенкевич. М.: Просвещение. 1968. С. 156-
193.
Алексеев В.В., Гирусов Э.В. Энергетика и проблемы экологии // Пущине:
Научный центр биологических исследований АН СССР. 1981 14 с.
(препринт)
Алексеева Т.А. Стандартный обмен ракообразных // Известия РАН. Сер.
биол. 1999. № . С.
Алексеева Т.А., Зотин А.И. Энергетический обмен у насекомых:
"прямокрылые, стрекозы, жуки // Известия РАН. Сер. биол. 1995. №3.
С.316-326.
Алексеева Т.А., Зотин А.И. Стандартный обмен насекомых: клопы, бабочки,
перепончатокрылые // Известия РАН. Сер.биол. 1996 №2. С. 193-205.
Алексеева Т.А., Зотин А.И., Донцова Г.В. Потребление кислорода
половозрелыми формами тараканов (Blattodea: Insecta) // Известия
РАН. Сер.биол. 1998. №5. С.632-636.
Алексеева Т.А., Зотин А.И., Ясюкевич В.В., Сидорова Н.В. Потребление
кислорода взрослыми формами двукрылых насекомых // Известия
РАН. Сер.биол. 1993. №3. С.426-435.
Алимов А.Ф. Интенсивность обмена у пресноводных двустворчатых
моллюсков // Экология. 1975. №1. С. 10-20.
Алимов А.Ф. Функциональная экология пресноводных двустворчатых
моллюсков. М.: Наука. 1981. 248 с.
Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Пятиязычный
словарь названий животных. Амфибии и рептилии. М.: Русский
язык. 1988. 560 с.
Аннинский Б.E. Энергетический баланс медузы Aurélia aurita L. в условиях
Черного моря // Биоэнергетика гидробионтов / Ред. Г.Е.Шульман,
Г.А.Финенко. Киев: Наукова Думка. 1990. С. 11-32.
АнриВ. Энергетика жизни//Природа. 1917. Т4. С.443-456.
Антонов В.Ф. Термодинамика биологических систем // Биофизика / Ред.
Б.Н.Тарусов, О.Р.Кольс. М.: Высшая школа. 1968. С.13-47.
Аринчин Н.И. Старение и продолжительность жизни животных в филогенезе
// Биология старения / Ред. В.В.Фролькис. Л.: Наука. 1982. С.39-60.
Аршавский И.А. Очерки по возрастной физиологии. М.: Медицина. 1967.
476 с.
Аршавский И.А. Физиологические механизмы и закономерности
индивидуального развития. М.: Наука. 1982. 270 с.
Ауэрбахъ Ф. Эктропизм или физическая теория жизни. СПб.: Образование,
1911. 114с.
Ауэрбах Ф. Основные понятия современного естествознания. М.: Фаросъ.
1913. 178с.
Афанасьев С.Л. Геохронологическая шкала фанерозоя и проблема
геологического времени. М.: Недра. 1987. 144 с.
Багдасарова Т.А. Исследование тканевого дыхания у памирской полевки
(Alucola argentatus) и у узкочерепной полевки (Microtus stenocranius)
II Температурная компенсация и поведенческий гомеостаз / Ред.
К.П.Иванов, А.Д.Слоним. Л.: Наука. 1980. С.45-52.
Базилевич Н.И., Родин Л.Е., Розов H.H. Сколько весит живое вещество
планеты? // Природа. 1971. № 1. С. 46-53.
Байченко И.П. Газообмен на разных высотах // Бюлл. Всесоюзн.НИИ эксп.
медицины. 1935. №2. С.31-32.
Банников А.Г., Даревский И.С., Денисова М.Н., Дроздов H.H. Класс
пресмыкающихся, или рептилий (Reptilia) // Жизнь животных.
Земноводные, пресмыкающиеся / Ред. А.Г.Банников. М.:
Посвящение. 1969. Т.4. Часть 2. С. 134-481.
Барбашова З.И. Акклиматизация к гипоксии и ее физиологический
механизм. М./Л.: Изд-во АН СССР. 1960. 216 с.
Барбашова З.И. Динамика повышения резистентности организма и
адаптивных реакций на клеточном уровне в процессе адаптации к
гипоксии // Успехи физиол. наук. 1970. Т.1. №3. С.70-88.
Бауэр Э.С. Теоретическая биология. М./Л.: Изд-во ВИЭМ, 1935. 206 с.
Бахарева И.Ф. Нелинейная неравновесная термодинамика. Саратов: Саратов.
ун-т. 1976. 140 с.
Бахем И., Лампрехт И. Гнездо лесных муравьев Formica polycena как модель
экологической системы // Журнал общей биологии. 1983. Т.44. № 1.
С.114-123.
Бей-Биенко Г.Я. Общая энтомология. М.: Высшая школа. 1980. 416 с.
Белокопытин Ю.С. Исследование основного обмена у черноморских рыб //
Тр-ды Всесоюзн. об-ва гидробиол. 1973. Т.18. С.117-128.
Белокопытин Ю.С. Изучение скорости потребления кислорода у кальмаров
тропической зоны Атлантического океана // Экология моря / Ред.
В.Е.Заика и др. Киев: Наукова Думка. 1982. №10. С.81-86.
Берг Л.С. Номогенез, или эволюция на основе закономерностей. Петербург:
Госиздат. 1922. 306 с.
Берг Л.С. Труды по теории эволюции. Л.: Наука. 1977. 387 с. Бергман Ю.Э.
Взаимозависимость интенсивности обмена и скорости роста у
морского ежа Strongylocentotus intermedius (Echinodermata,
Strongylocentrotida.). Зоол. журнал. 1971. Т.50, №10. С. 1530-1538.
Беркинблит М.Б. О применимости теоремы Пригожина к процессу
эмбриогенеза и эволюции // Биофизика. 1965. Т. 10, №6. С. 1103-1105
Беркович E.M. Энергетический обмен в норме и патологии. М.: Медицина.
1964. 333 с.
Бирштейн Я.А. Подтип Branchiata // Жтзнь животных. Беспозвоночные. /
Ред. Зенкевич Л.А. М.: Просвещение. 1968. Т.2. С.377-529.
Благовещенский A.B. Биологическая эволюция растений в связи с
изменением качества ферментов // Успехи совр. биологии. 1939. Т.11.
№3. С.320-339.
Благовещенский A.B. Биохимические основы эволюционного процесса у
растений. М.-Л.: Изд-во АН СССР. 1950. 271 с.
Блинков С.М. Количественные определения морфологических структур
центральной нервной системы // Итоги науки и техники. Морфология
человека и животных / Ред. В.Д.Захаренко, К.В.Савич. М.; ВИНИТИ.
1972. Т.5. С.5-141.
Блюменфельд Л.А. Проблемы биологической физики. М.: Наука. 1977. 336 с.
Больцман Л. Статьи и речи. М.: Наука. 1970. 406 с.
Борисов А.П. Предельное потребление кислорода при плавании //
Исследование по физиологии выносливости. М.; Л.: Физкультура и
спорт. 1949. С. 113-124.
Бриллюэн Л. Наука и теория информации. М.: Физматгиз. 1960. 392 с.
Бриллюэн Л. Термодинамика, статистика и информация // Успехи физ. наук.
1962. Т.77. №2. С.337-352.
Брода Э. Эволюция биоэнергетических процессов. М.: Мир. 1978. 304 с.
Брэм А.Э. Жизнь животных. Т.3. М.- Л.: Молодая Гвардия. 1930. 704 с.
Брэм А.Э. Жизнь животных. Т.1. М.- Л.: Молодая Гвардия. 1931. 608 с.
Будыко М.И. Эволюция биосферы. Л.: Гидрометиоиздат. 1984. 488 с.
Бызова Ю.Б. Дыхание почвенных беспозвоночных // Экология почвенных
беспозвоночных/ Ред. М.С.Гиляров. М.: Наука. 1972. С.3-39.
Быстрай Г.П. Теплофизика. Термодинамика необратимых процессов.
Свердловск: Изд-во Уральского ун-та. 1987. 99 с.
Быховский А.И. Еще к вопросу о применимости теоремы Пригожина в
биологии (о повышении термогенеза на начальных стадиях развития
живых организмов) // Биофизика. 1965. Т. 10. №6. С. 1105-1108.
Быховский А.И. О тепловых эффектах и надежности процессов синтеза
белков и нуклеиновых кислот // Доклады АН СССР. 1966. Т. 171, №4.
С.992-995.
Вассоевич Н.Б., Иванов А.Н. К истории учения о биосфере // Методология и
история геологических наук / Ред.А.И.Равикович и др. М.: Наука.
1977. С.57-94.
Вассоевич Н.Б., Иванов А.Н. О различном понимании биосферы //
Палеонтология и эволюция биосферы / Ред.А.Н.Олейников и др. Л.:
Наука. 1983. С. 17-33.
Верзилин H.H., Верзилин H.H., Верзилин Н.М. Биосфера, ее настоящее,
прошлое, будущее. М.: Просвещение. 1976. 223 с.
Вернадский В.И. Биосфера. Л.: Научи, хим. техн. изд-во. 1926. 146 с.
Вернадский В.И. Биогеохимические очерки. М./Л.: Изд. АН СССР. 1940.250с.
Вернадский В.И. Биосфера. М.: Мысль. 1967. 374 с.
Виленкина М.Н. Соотношение дыхания Actinia equina L. на тканевом и
организменном уровнях // Физиология морских животных. Биология
моря. М.: Наука. 1966. С.233-245.
Виленкина М.Н., Виленкин Б.Я. О возможности функционального подхода к
количественной оценке степени дифференциации и интеграции
организма // Журнал общей биологии. 1969. Т.30. №2. С. 132-139.
Винберг Г.Г. Интенсивность обмена у простейших // Успехи совр. биологии.
1949. Т.28. №2(5). С.226-245.
Винберг Г.Г. Интенсивность обмена и размеры ракообразных // Журнал
общей биологии. 1950. Т.Н. №5. С.367-380.
Винберг Г.Г. Интенсивность обмена и пищевые потребности рыб. Минск:
Изд-во Белорусск. ун-та. 1956. 253 с.
Винберг Г.Г. Зависимость энергетического обмена от массы тела у водных
пойкилотермных животных // Журнал общей биологии. 1976. Т.37.
№1. С.56-70.
Виноградов А.П. Химический элементарный состав организмов моря. Часть
I // Труды биогеохимической, лаборатории. 1932. Т.2. С.5-122.
Виноградов А.П. Химический элементарный состав организмов моря. Часть
II // Труды биогеохимической, лаборатории. 1937. Т.4. С.5-225.
Виноградов А.П. Химический элементарный состав организмов моря. Часть
III // Труды биогеохимической лаборатории. 1944. Т.6. С.5-273.
Виноградова Н.Г. Тип Оболочники (Tunicata) // Жизнь животных.
Беспозвоночные (Ред. Л.А.Зекнкевич). М.: Просвещение. Т.2. С.329-
363.
Владимирова И.Г. Дыхание при регенерации и заживлении раны у планарии
Dendrocoelum lacteum II Известия АН СССР. Сер.биол. 1980. №5.
С.781-785.
Владимирова И.Г. Макросистематика брюхоногих моллюсков по
энергетическрму обмену Известия АН СССР. Сер.биол. 1999. № .
С.
Владимирова И.Г., Алексеева Т.А., Никольская И.С., Радзинская Л.И., Зотин
А.И. Интенсивность потребления кислорода и процесс неотении у
амбистом // Журнал общей биологии. 1993. Т.54. №1. С.45-49.
Владимирова И.Г., Донцова Г.В. Влияние кормления и голодания на
дыхание и вес тела планарии // Известия АН СССР. Сер.биол. 1979.
№6. С.922-926.
Владимирова И.Г., Зотин А.И. Скорость дыхания у простейших. М., 1983.
Деп. в ВИНИТИ. №5500-83. 61 с.
Владимирова И.Г., Зотин А.И. Зависимость скорости дыхания простейших
от температуры и веса тела // Журнал общей биологии. 1985. Т.46.
№2. С. 165-173.
Владимирова И.Г., Зотин А.И. Данные о влиянии температуры на
потребление кислорода у животных. 1. Млекопитающие. Деп.
ВИНИТИ. 1986. №3659-В86. 85.
Владимирова И.Г., Зотин А.И. Кривые терморегуляции и определяющие их
факторы//Успехи физиол. наук. 1989. Т.20. №3. С.21-42.
Владимирова И.Г., Зотин А.И. Стандартный обмен в классе амфибий //
Известия РАН. Сер.биол. 1994. №1. 81-92.
Владимирова И.Г., Сербинова И.А. Зависимость интенсивности дыхания от
массы тела у некоторых представителей хвостатых и бесхвостых
земноводных // Журнал общей биологии. 1992. Т.53. №5. С.744-749.
Властов Б.В., Матекин П.В. Класс Панцирных (Loricata) // Жизнь животных /
Ред.Л.А.Зенкевич. М.: Наука. 1968. Т.2. С. 14-19.
Водопьянов H.A. Устойчивость и динамика биосферы. Минск: Наука и
техника. 1981. 246 с.
Волькенштейн М.В. Общая биофизика. М.: Наука. 1978. 590 с.
Волькенштейн М.В., Сырников Ю.П. Еще раз об "Элементах биологической
термодинамики" К.С.Тринчера // Биофизика. 1966. Т.Н. №2. С.379-
380.
Воробьева Э.И. Проблема происхождения наземных позвоночных. М.:
Наука. 1992. 344 с.
Воронин Н.М., Спиридонова Ф.В., Новиков М.Н. Изменение степени
насыщения крови кислородом, газообмен и корковые реакции у
больных туберкулезом легких в период акклиматизации к
высокогорному климату курорта "Цей" // Проблемы
экспериментальной курортологии./Ред. Ф.Д.Васильев. М.: Медгиз.
1958. Т.З. С.83-85.
Воронцов H.H. Синтетическая теория эволюции: ее источники, основные
постулаты и нерешенные проблемы // Журнал Всесоюзн. хим. об-ва
им Д.И.Менделеева. 1980. Т.25. №3. С.293-312.
Воронцов H.H. Теория эволюции: истоки, постулаты и проблемы. М.:
Знание. 1984. 64 с.
Воронцова М.А., Лиознер Л.Д.„ Маркелова Л.В., Пучальская Б.Ч. Тритон и
аксолотль. М.: Советская наука. 1952. 295 с.
Гаврилов В.В. Максимальная продолжительность жизни свободноживущих
куликов - зависимость от массы // Известия РАН, Сер. биол. 1991.
№3. С.471-473.
Гаврилов В.В., Зотин А.И. Данные о стандартном обмене птиц. База данных
Лаборатории биофизики развития Института биологии развития
РАН. (340 видов). 1998.
Гаврилов В.М. Направление биоэнергетических адаптации у птиц к
сезонности климата // Экология, география и охрана птиц. / Ред.
И.А.Нейфельдт. Л.: Зоол. ин-т. 1980. С.73-97.
Галл Я.М. Единство механизмов микро- и макроэволюции // Развитие
эволюционной теории в СССР / Ред. С.П.Микулинский,
Ю.И.Полянский. Л.: Наука. 1983. С.259-262.
Гандельсман А.Б., Васильева В.В. Показатели состояния тренированности //
Физиология мышечной деятельности труда и спорта / Ред.
К.М.Смирнов. Л.: 1969. С.393- 401.
Геккер Р.Ф. (Ред.) Основы палеонтологии. Справочник для палеонтологов и
геологов СССР. Т. 10. Иглокожие, гемихордовые. М.: Недра. 1964.
383с.
Гельфер Я.M. История и методология термодинамики и статистической
физики. T.I. M.: Высшая школа. 1969. 475 с.
Гельфер Я.М. История и методология термодинамики и статистической
физики. Т.2. М.: Высшая школа. 1973. 280 с.
Гептнер В.Г., Насимович A.A., Банников А.Г. Млекопитающие Советского
Союза. Т.1. Парнокопытные и непарнокопытные. М.: Высшая школа.
1961.776с.
Гиляров М.С. Тип Членистоногие (Arthropoda). Введение. // Жизнь
животных / Ред. Л.А.Зенкевич. 1968. Т.2. С.367-376.
Гладышев Г.П. О термодинамике биологической эволюции. Черноголовка:
Ин-т хим.-физики АН СССР. 1977. 46 с.
Гладышев Г.П. Термодинамика и макрокинетика природных иерархических
процессов. М.: Наука. 1988. 287 с.
Гладышев Г.П. Термодинамическая теория эволюции живых существ. М.:
Луч. 1996. 86с.
Гладышев Г.П., Гладышев Д.П. О модели эволюции биологических систем //
Известия РАН. Сер.биол. 1994. №1. 14-19.
Глене дорф П., Пригожин И. Термодинамическая теория структуры,
устойчивости и флуктуации. М.: Мир. 1973. 280 с.
Голубев B.C. Модель эволюции геосфер. М.: Наука. 1990. 95 с.
Голубев B.C. Термодинамический критерий эволюции и развитие биосистем
//Журн. общ. биологии. 1991. Т.52. №2. С.149-161.
Горшков В.Г. Энергетика биосферы и устойчивость состояния окружающей
среды // Итоги науки и техники. Сер. Теоретические и общие
вопросы географии / Ред.К.С.Лосев и др. М.: ВИНИТИ. 1990. Т.7.
С.3-238.
Грант В. Эволюция организмов. М.: Мир. 1980. 480 с.
Грант В. Эволюционный процесс: Критический обзор эволюционной теории.
М.:Мир. 1991.488с.
Громова В.И. (Ред.) Основы палеонтологии. Справочник для палеонтологов
и геологов СССР. Т. 13. Млекопитающие. М.: Гос. научно-техн. изд-
во литературы по геологии и охране недр . 1962. 421 с.
Гуров К.П. Феноменологическая термодинамика необратимых процессов.
М.: Наука. 1978. 128с.
Давиташвили Л.Ш. Очерки по истории учения об эволюционном прогрессе.
М.: Изд-во АН СССР. 1956. 228 с.
Давиташвили Л.Ш. Учение об эволюционном прогрессе (теория
ароморфоза). Тбилиси: Медниуруба. 1972. 324 с.
Дарвин Ч. Происхождение видов путем естественного отбора. М./Л. : Изд-во
АН СССР. 1939. Собр. соч. Т.З, 589 с.
Де Гроот С. Термодинамика необратимых процессов. М.: Гос. изд-во техн.-
теорет. литературы. 1956. 280 с.
Де Гроот С., Мазур П. Неравновесная термодинамика. М.: Мир. 1964. 456 с.
Демин В.И., Андросова И.М., Озернюк Н.Д. Адаптация энергетического
обмена у рыб: Влияние скорости плаванья и температуры на
цитохромную систему скелетных мышц // Докл. АН СССР. 1989.
Т.308. №1. С.241-246.
Денбиг Л. Термодинамика стационарных необратимых процессов. М.: Изд-во иностр. Литературы. 1954. 119 с.
Джоунс М. (Ред.) Биохимическая термодинамика. М.: Мир. 1982. 440 с.
Догель В.А. Зоология беспозвоночных. М.: Высшая школа. 1947. 527 с.
Дольник В.Р. Энергетический обмен и эволюция животных // Успехи
соврем, биологии. 1968. Т.66. №5. С.276-293.
Дольник В.Р. Энергетический метаболизм и размеры животных: Физические
основы соотношения между ними // Журнал общ.биологии. 1978.
Т.39. №6. С.805-816.
Дольник В. Р. Аллометрия морфологии, функции и энергетики
гомойотермных животных и ее физический контроль // Журнал общ.
биологии. 1982. Т.43. №4. С.435-454.
Донцова Г.В., Алексеева Т.А., Зотин А.И. Зависимость интенсивности
дыхания тараканов от веса тела и температуры // Известия АН СССР.
Сер. биол. 1985. №5. С.787-789.
Донцова Г.В., Зотин А.И. Зависимость максимального дыхания животных от
веса тела и уровня стандартного обмена // Термодинамика и кинетика
биологических процессов / Ред. А.И.Зотин. М.: Наука. 1980. С.282-
294.
Дуршиц В.В., Обручева О.П. Палеонтология. М.: Изд-во Московского Гос.
Университета. 1971. 414 с.
Дьярмати И. Неравновесная термодинамика. Теория поля и вариационные
принципы. М.: Мир. 1974. 304 с.
Ершов Ю.А. Термодинамика квазиравновесий в биологических системах //
Итоги науки и техники. Химическая термодинамика и равновесия.
/Ред. А.Н.Саприн. М.: ВИНИТИ. 1983. Т.5. 140 с.
Ефимов Г.О. Предельное потребление кислорода при беге на месте у лиц с
различной степенью тренировнности // Исследования по физиологии
выносливости / Ред. С.В.Фарфуль. М./Л: Физкультура и спорт. 1949.
С.125-152.
Жакин A.B. Сезонные изменения интенсивности обмена у дальневосточного
трепанга Stichopus japonicus в заливе Посьета (Японское море) //
Извучтия. ТИНРО. 1982. Т. 106. С. 146-148.
Желявский В., Галинский В. Термодинамические аспекты онтогенеза
растений // Термодинамика и регуляция биологических процессов /
Ред. А.И.Зотин. М.: Наука. 1984. С.302-308.
Журавлев В.А. Термодинамика необратимых процессов в задачах и
решениях. М.: Наука. 1979. 135 с.
Завадский K.M. К пониманию прогресса в органической природе //
Проблемы развития в природе и обществе / Ред. Б.А.Чагин. М.-Л.:
Изд-во АН СССР. 1958. С.79-120.
Завадский K.M. Движущая сила арогенеза // Проблемы прогрессивного
развития в природе и технике / Ред. И.Г.Васильев и др. Л.: Наука.
1969. С.64-69.
Завадский K.M. К исследованию движущих сил арогенеза // Журнал общ.
биологии. 1971. Т.32. №5. С.515-529.
Завадский K.M. Развитие эволюционной теории после Дарвина. Л.: Наука.
1973. 423 с.
Завадский K.M., Ермоленко М.Т. К критике неономогенеза // Философские
проблемы современной биологии / Ред. А.С.Мамзин. М.-Л.: Наука.
1966. С.227-233.
Завадский K.M., Жердев Р.В. Проблема специализации в эволюционной
теории // Философские проблемы эволюционной теории / Ред.
Б.Е.Быховский и др. М.: Наука. 1971. С.55-68.
Завадский K.M., Колчинский Э.И. Эволюция эволюции. Л.: Наука. 1977.236с.
Завальнюк П.А. Термодинамическая теория вероятностных распределений и
управление качеством Ч. 1 Тверь: Тверской гос. ун-т. 1991. 75 с.
Залевски К. Феноменологическая и статистическая термодинамика.
М.:Мир. 1973. 167с.
Зарафянц М.Т. Что такое энтропия. Харьков: Гостехиздат Украины. 1935.84с.
Захаров В.M. Гомеорез: онтогенетический, популяционный и эволюционный
анализ // Термодинамика и регуляция биологических процессов / Ред.
А.И.Зотин. М.: Наука. 1984. С.294-302.
Захаров В.М. Асимметрия животных. М.: Наука. 1987. 216 с.
Зотин A.A. Статистическая оценка параметров аллометрический уравнений
// Известия РАН. Сер. биол. 1999. № . С. - .
Зотин A.A., Алексеева Т.А., Владимирова И.Г., Зотин А.И. Эволюционная
биоэнергетическая ловушка и прогрессивная эволюция животных //
Известия РАН. Сер. биол. 2000 (в печати)
Зотин A.A., Алексеева Т.А., Зотин А.И. Стандартный обмен паукообразных
// Известия РАН. Сер. биол. 1998а. №6. С.686-693.
Зотин A.A., Лампрехт И., Зотин А.И. Тепловые барьеры в процессе
прогрессивной эволюции животных и человека // Известия РАН.
Сер.биол. 19986. №3. С.305-315.
Зотин А.И. Изменение скорости продукции энтропии во время
эмбрионального развития и роста // Биофизика. 1966. Т.Н. №3.
С.554-557.
Зотин А.И. По поводу использования понятий энтропия и негэнтропия в
биологии//Журнал общ. биологии. 1971. Т.32. №1. С.27-36.
Зотин А.И. Термодинамический подход к проблемам развития, роста и
старения. М.: Наука. 1974. 183 с.
Зотин А.И. (Ред.) Термодинамика биологических процессов. М.: Наука. 1976.
278с.
Зотин А.И. Классификация птиц по энергетическому метаболизму // Aktualne
problmy aviarnej genetiky. Smolenice: Slovensk. Акад. 1979. S.33-55.
Зотин А.И. (ред.) Термодинамика и кинетика биологических процессов. М.:
Наука. 1980. 397 с.
Зотин А.И. (ред.) Термодинамика и регуляция биологических процессов. М.:
Наука. 1984. 334 с.
Зотин А.И. Биоэнергетическая направленность эволюционного прогресса
организмов // Термодинамика и регуляция биологических процессов /
Ред.А.И.Зотин. М.: Наука. 1984. С.269-274.
Зотин А.И. Термодинамическая основа реакций организмов на внешние и
внутренние факторы. М.: Наука. 1988. 272 с.
Зотин А.И. Прогрессивная эволюция животных. 1. Константа Рубнера в
классе млекопитающих // М.: Деп. ВИНИТИ. 1993а. №762-В93.
Зотин А.И. Прогрессивная эволюция животных. 2. Коэффициент
энцефализации и продолжительность жизни в классе
млекопитающих//М.: Деп. ВИНИТИ. 19936. №763-В93.
Зотин А.И. Данные о стандартном обмене рептилий // База данных
Лаборатории биофизики развияия Института биологии развития РАН
. (188 видов). 1998.
Зотин А.И., Алексеева Т.А. Надежность и устойчивость онтогенезов //
Термодинамика и регуляция биологических процессов / Ред.
А.И.Зотин. М.: Наука. 1984. С.288-294.
Зотин А.И., Алексеева Т.А. Прогрессивная эволюция и макросистематика в
классе насекомые // Изв. РАН. Сер. биол. 1997. №2. С. 186-195.
Зотин А.И., Владимирова И.Г. Данные о стандартном обмене животных. 1.
Млекопитающие: все отряды кроме шерстокрылых, трубкозубых и
грызунов. Деп.ВИНИТИ. 1986а. №3660-В86. 70 с.
Зотин А.И., Владимирова И.Г. Данные о стандартном обмене животных. 2.
Млекопитающие: грызуны. Деп.ВИНИТИ. 19866. №5903-В86. 70 с.
Зотин А.И., Владимирова И.Г. Макросистематика млекопитающих по
критериям энергетического метаболизма // Известия АН СССР.
Сер.биол. 1991. №1. С.59-69.
Зотин А.И., Владимирова И.Г. Данные о стандартном обмене животных. 5.
Амфибии. Деп.ВИНИТИ. 1993; №764-В93. 34 с.
Зотин А.И., Владимирова И.Г., Кирпичников A.A. Энергетический
метаболизм и направление эволюционного прогресса в классе
млекопитающих // Журнал общ. биологии. 1990. Т.51. №6. С.760-767.
Зотин А.И., Зотин A.A. Прогрессивная эволюция: термодинамическая основа
// Известия РАН. Сер.биол. 1995. №4. С.389-397.
Зотин А.И., Зотин A.A. Данные о стандартном обмене моллюсков:
двустворчатые. База данных Лаборатории биофизики развития
Института биологии развития РАН. (50 видов). 1998.
Зотин А.И., Зотин A.A. Данные о стандартном обмене рыб. База данных
Лаборатории биофизики развития Института биологии развития
РАН. (236 видов). 1999.
Зотин А.И., Зотина P.C. Экспериментальная и теоретическая основа
качественной феноменологической теории развития //
Термодинамика биологических процессов / Ред. А.И.Зотин М.:
Наука. 1976. С.47-65.
Зотин А.И., Зотина P.C. Функция внешней диссипации и эволюция открытой
системы к стационарному состоянию // Теоретическая и
экспериментальная биофизика / Ред. А.К.Приц. Калининград. 1977.
Вып.7. С.10-18.
Зотин А.И., Зотина P.C. Термодинамические критерии устойчивости и
надежности биологических систем и процессов развития //
Надежность и гомеостаз биологических систем / Ред.
.М.Гродзинский. Киев: Наукова Думка. 1987. С.26-34.
Зотин А.И., Зотина P.C. Феноменологическая теория развития, роста и
старения организмов. М.: Наука. 1993. 364 с.
Зотин Α., Зотина Р. Освоение космоса - императив выживания человека. //
Здравый смысл. 1998. № 9. С. 47-49.
Зотин А.И., Коноплев В.А. Направление эволюционного прогресса
организмов // Термодинамика биологических процессов / Ред.
А.И.Зотин. М: Наука. 1976. С.230-235.
Зотин А.И., Коноплев В.А., Радзинская Л.И., Никольская И.С. Зависимость
скорости потребления кислорода от массы тела мидий // Гидробиол.
журн. 1987. Т.23. №2. С.64-67.
Зотин А.И., Криволуцкий Д.А. Скорость и направление эволюционного
прогресса организмов // Журнал общ. биологии. 1982. Т.43. №1. С.З-
13.
Зотина P.C., Зотин А.И. Кинетика конститутивных процессов во время
развития и роста организмов // Термодинамика и кинетика
биологических процессов / Ред. А.И.Зотин. М.: Наука. 1980. С.318-
327.
Ибраимова Г.И. Акклимация (акклиматизация) и температурная
компенсация у пойкилотермных организмов // Экологическая
физиология животных / Ред. А.Д.Слоним. Л.: Наука. 1982. Ч.З. С.24-
41.
Иванов К.П. Гомойотермия и энергетика гомойотермного организма //.
Физиология терморегуляции / Ред. К.П.Иванов и др.. Л.: Наука. 1984.
С.7-28.
Иванов К.П. Основы энергетики организмов. Т.1. Общая энергетика,
теплообмен и терморегуляция. Л.: Наука. 1990. 307 с.
Ив лев B.C. Зависимрсть интенсивности обмена у рыб от веса их тела //
Физиологический журн. 1954. Т.40. №6. 717-721.
Ив лев В. С. Опыт оценки эволюционного значения уровней энергетического
обмена // Журнал общ. биологии. 1959. Т.20. №6. С.94-103.
Ив лев B.C. Уровень энергетического обмена ланцетника // Доклады АН
СССР. 1961. Т. 140. №5. С. 1217-1219.
Ив лев B.C. О параметрах, характеризующих уровни энергетического обмена
животных // Применение математических методов в биологии / Ред.
Л.С.Каминский, В.П.Терентьев. Л.: Изд-во ЛГУю 1963. Т.2. С. 146-
151.
Ив лев B.C., Яковлева К. К. Уровень энергетического обмена губок //
Доклады АН СССР. 1963. Т. 152. №1. С.241-243.
Ивлева И.В. Элементы энергетического баланса актиний // Труды
Севастопольской биол. станции. 1964. Т. 15. С.410-428.
Ивлева И.В. Влияние температуры на скорость метаболизма
пойкилотермных животных//Успехи совр. биологии. 1972. Т.73. №1.
С.134-155.
Ивлева И.В. Температура среды и скорость энергетического обмена у
водных животных. Киев: Наукова думкаю 1981. 232 с.
Ивлева И.В., Литовченко Т.И. Скорость дыхания Coelenterata и Ctenophora в
зависимости от температуры среды обитания // Биология моря. Киев.
1978. №46, С.3-25.
Ивлева И.В., Попенкина М.И. О температурной зависимости обмена у
пойкилотермных животных // Физиологические основы экологии
водных животных. Биология моря. !968. Т. 15 С.29-51.
Иорданский H.H. Неравномерность темпов макроэволюции и ключевые
ароморфозы // Природа. 1977. №6. С.36-36.
Иорданский H.H. Основы теории эволюции. М.: Наука. 1979. 190 с.
Иорданский H.H. Эволюция комплексных адаптации. Челюстной аппарат
амфибий и рептилий. М.: Наука. 1990. 310 с.
Иорданский H.H. Микроэволюция: системная теория. М.: Наука. 1994. 112 с.
Калабухов Н.И. Спячка млекопитающих. М.: Наука. 1985. 260 с.
Калабухов H.И., Ладыгина H.M. Возникновение эколого-физиологических
особенностей у млекопитающих под воздействием внешней среды //
Зоол. журнал. 1953. Т.32. №2. С.294-299.
Камлюк Л.В. Энергетический обмен у свободноживущих плоских и
кольчатых червей и факторы, его определяющие // Журнал общ.
биологии. 1974. Т.35. №6. С.874-885.
Камлюк Л.В., Кожова О.М., Кукушкина Л.М. Об уровне энергетического
обмена у планарий отряда Triclada (Plathelminthes, Turbellaria) //
Доклады АН Белорусской ССР. 1974. Т. 18. №4. С.376-378.
Камшилов М.М. Значение взаимных отношений между организмами в
эволюции. М./Л.: Наука. 1961. 136 с.
Камшилов М.М. Эволюция организованности // Философские проблемы
эволюционной теории / Ред. Б.Е.Быховский и др. М.: Наука. 1971.
Т.1.С.ЗО-37.
Камшилов М.М. Эволюция биосферы. М.: Наука. 1979. 256 с.
Капанадзе И.С., Капанадзе Б.И. Длительность онтогенетического цикла у
растений некоторых родов подсемейства померанцевых //
Онтогенез. 1995. Т.26. №4. С.293-299.
Карташев H.H. Систематика птиц. М.: Высшая школа. 1974. 367 с.
Кашкаров Д.Н., Станчинский В.В. Курс зоологии позвоночных. М.: Изд-во
АН СССР. 1940. 1024 с.
Кеплен С. Р., Эссиг Э. Биоэнергетика и линейная термодинамика
необратимых процессов. М.: Мир. 1986. 382 с.
Клаузиус Р. Механическая теория теплоты // Второе начало термодинамики /
Ред. А.К.Тимирязев. М./Л.: Гостехиздат. 1934. С.73-158.
Кнорре Е. Наука плюс фантазия. М.: Детская литература. 1978. С.8-9.
Ковальский В.В. Геохимическая экология. М.: Наука. 1974. 299 с.
Ковальский В.В. Геохимическая среда и жизнь. М.: Наука. 1982. 77 с.
Ковда В.Α., Янушевская И.В. Опыт оценки биомассы суши // Известия АН
СССР. Сер.биол. 1967. № з. С.ЗЗ 1-338.
Колчинский Э.И. Эволюция биосферы. Л.: Наука. 1990. 236 с.
Коноплев В.А., Зотин А.И. Критерий упорядоченности и его применение к
некоторым эволюционным проблемам // Теоретическая и
экспериментальная биофизика./ Ред. А.К.Приц. Калининград. 1975.
Вып.5. С.98-108.
Коноплев В.А., Соколов В.Е., Зотин А.И. Классификация животных по
энергетическому метаболизму // Зоол. журнал. 1975. Т.54. №10.
С. 1429-1441.
Коноплев В.А., Соколов В.Е., Зотин А.И. Критерий упорядоченности и
проблемы классификации животных // Термодинамика
биологических процессов / Ред. А.И.Зотин. М: Наука. 1976. С.235-
243.
Кордюм В.А. Эволюция и биосфера. Киев: Наукова Думка. 1982. 264 с.
Корнишъ Ч.Д. Жизнь животных в фотографиях с натуры. М.: Кнебель. 1907.
383с.
Корочкин Л.И. К спорам о дарвинизме // Химия и жизнь. 1982. №5. С.57-61.
Корочкин Л.И. К вопросу о направленности молекулярных и
форообразовательных событий в онто- и филогенезе // Молекулярная
генетика и биофизика / Ред. Н.Е.Кучеренко и др. Киев: Вища школа.
1985. С.82-95.
Корочкин Л.И., Ивановский А.Б. Скачки в эволюции // Химия и жизнь. 1983.
№10. С.40-47.
Костицын В.А. Эволюция атмосферы, биосферы и климата. М.: Наука. 1984.
96с.
Крепе Е.М., Вержбинская H.A., Ченыкаева Е.Ю., Чирковская Е.В.,
Гавырина Ц.К. О приспособлении животных к хронической гипоксии //
Физиол. журнал СССР. 1956. Т.42. №1. С.69-77.
Кривошеев В.Г., Бурмакин В.Н. Изменчивость адаптивных
морфофизиологических признаков полевок с широкими и узкими
ареалами // Биологические проблемы Севера. / Ред.
В.Л.Комтримавичус. Магадан. 1971. С.5-35.
Крушинская Н.Л., Лисицына Т.Ю. Поведение морских млекопитающих. М.:
Наука. 1983. 336 с.
Крушинский Л.В. Биологические основы рассудочной деятельности:
Эволюционные и физиолого-генетические основы поведения. М.:
Изд-во МГУ. 1977. 270 с.
Кузьмичова В.И. Уровень дыхания медуз Черного моря // Экосистемы
пелагиали Черного моря / Ред. М.Е.Виноградов. М.: Наука. 1980.
С.219-223.
Кэрролл Р. Палеонтология и эволюция позвоночных. Т1. М.: Мир. 1992.280с.
Кэрролл Р. Палеонтология и эволюция позвоночных, т.2. М.: Мир. 1993а.
283с.
Кэрролл Р. Палеонтология и эволюция позвоночных. т.З. М.: Мир. 19936.
312с.
Лазарева Л.П. О потреблении кислорода гребневиками Pleurobrachia pileus
O.F.Mller разных размеров в зависимости от температуры и
солености окружающей среды // Тр-ды Карадагской биол. станции.
1961.Т.17. С.85-96.
Лазарев П.П. Современные проблемы биофизики. М.-Л.: Изд-во АН СССР.
1945. 152 с.
Ламарк Ж. Философия зоологии. М.-Л.: Гос. изд-во биол. мед. литерат.,
1935.1.1.330с.
Ламарк Ж. Философия зоологии. М.-Л.: Гос. изд-во биол. мед. литерах.,
1937. Т.2. 483 с.
Лампрехт И. Энтропия и информация - сверхъестественное поведение
демона Максвелла // Термодинамика и регуляция биологических
процессов / Ред. А.И.Зотин. М.: Наука. 1984. С.82-97.
Ланге А. Б. Подтип Трилобиты (Trilobita) // Жизнь животных.
Беспозвоночные / Ред.Л.А.Зенкевич. М.: Просвещение. 1969а. Т.З.
С.7-9.
Ланге А. Б. Подтип Хелицеровые (Chelicerata) // Жизнь животных.
Беспозвоночные / Ред.Л.А.Зенкевич. М.: Просвещение. 19696. Т.З.
С.14-17.
ЛевонтинР. Генетические основы эволюции. М.: Мир. 1978. 351 с.
Ливчак Г.Б. Материалы к сравнению характеристик дыхания тканей и
органов грызунов // Известия СО АН СССР. Сер.биол. 1972. Вып.З.
87-94.
Ломов И.А. О законе поверхности Рубнера // Количественные аспекты роста
организмов / Ред. А.И.Зотин, Г.А.Клевезаль, В.А. Грудницкий. М.:
Наука. 1975а. С. 181-184.
Ломов И.А. Относительная мощность и основной обмен // Количественные
аспекты роста организмов / Ред. А.И.Зотин Г.А.Клевезаль,
В.А.ГрудницкийМ.: Наука. 19756. С.244-248.
Лукашев К.И., Вадковская И.К. Биосфера и биогеохимические провинции.
Минск: Наука и техника. 1973. 239 с.
Лункевич В.В. От Гераклита до Дарвина. М.-Л.: Изд-во АН СССР. 1936. Т.1.
414с.
Лункевич В.В. От Гераклита до Дарвина. М.-Л.: Изд-во АН СССР. 1940. Т.2.
495с.
Лункевич В.В. От Гераклита до Дарвина. М.-Л.: Изд-во АН СССР. 1943. Т.З.
467с.
Луппов Н.П., Друщиц В.В. (Ред.) Основы палеонтологии. Справочник для
палеонтологов и геологов СССР. Т.6. Моллюски - головоногие, II:
аммоноидеи (цератиты, аммониты), внутрираковинные. Приложение
- кониконхии. М.: Гос. научно-техн. изд-во литературы по геологии и
охране недр. 1958. 359 с.
Ляпунов A.A. Об управляющих системах живой природы / О сущности
жизни/Ред. Г.М.Франк, А.М.Кузин. М.: Наука. 1964. С.66-80.
Майер Р. Закон сохранения и превращения энергии. М.: ГТТИ. 1933.
Майр Э. Зоологический вид и эволюция. М.: Мир. 1968.597 с.
Майр Э. Популяции, виды, эволюция. М.: Мир. 1974. 460 с.
Маргелис Л. Роль симбиоза в эволюции клетки. М.: Мир. 1983. 352 с.
Маршалл Э. Биофизическая химия. М.: Мир. 1981. Т.1. 358 с.
Матвеев B.C. (Ред.) Курс зоологии. Т.2. Хордовые. Изд.4. М.: Советская
наука. 1949. 519с.
Матвеев B.C. (Ред.) Курс зоологии. Т.2. Хордовые. М.: Советская наука. 1956.
444с.
Медников Б.М. Дарвинизм в XX веке. М.: Знание. 1973. 64 с.
Медников Б.М. Аксиомы биологии. Biologia axiomatica. M.: Мир. 1982. 136 с.
Медников Б.М. Об основных принципах теоретической биологии // Журнал
общ. биологии. 1984. Т.45. №6. С.723-731.
Медников Б.М. Н.В.Тимофеев-Ресовский и аксиоматика теоретической
биологии // Онтогенез. Эволюция. Биосфера. / Ред. А.В.Яблоков. М.:
Наука. 1989. С. 15-30.
Меерсон Ф.З. Общий механизм адаптации и профилактики. М.: Медицина.
1973. 360 с.
Меерсон Ф.З. Механизмы адаптации к высотной гипоксии // Итоги науки и
техники. Физиология человека и животных. Т. 14. Проблемы
гипоксии и гипероксии / Ред. Г.А.Степанский.. М.: ВИНИТИ. 1974.
С.7-62.
Меерсон Ф.З.Адаптация, стресс и профилактика. М.: Наука. 1981. 274 с.
Мейен C.B. Проблема направленности эволюции // Итоги науки и техники.
Зоология позвоночных. Проблемы теории эволюции / Ред.
Л.П.Познанин, А.В.Яблоков. М.: ВИНИТИ. 1975. Т.7. С.66-117.
Мейен C.B. Может ли быть победитель в дискуссии о номогенезе // Природа.
1979. №Ю. С. 114-116.
Мещеряков A.C., Улыбин С.А. Термодинамика. Феноменологическая
термомеханика. М.: Химия. 1994. 352 с.
Миклин A.M. Информационный критерий высоты организации и
прогрессивного развития живого // Проблемы прогрессивного
развития в живой природе и технике / Ред. И.Г.Васильев и др. Л.:
Наука. 1969. С.82-86.
Миклин A.M. Критерии прогрессивной эволюции // Развитие эволюционной
теории в СССР / Ред. С.П.Микулинский, Ю.И.Полянский. Л.: Наука.
1983. С.358-364.
Мильман Л.С. Гликолиз в эмбриональном развитии вьюна Misgurnus fossilis
II Журнал общ. биологии. 1965. Т.26. №2. С.237-240.
Мирзонян Э.Н. Учение А.Н.Северцова о главных направлениях
эволюционного процесса // Развитие эволюционной теории в СССР /
Ред. С.Р.Микулинский, Ю.П.Полянский. Л.: Наука. 1983. С.348-358.
Миррахимов Μ.,Μ. Основной обмен и его изменения в условиях
высокогорья // Газообмен в условиях высокогорья (в норме и
патологии) / Ред. Н.Н.Сиротинский.. Фрунзе: Кыргыстан. 1967. С.89-
106.
Михайлов В.В. Основной обмен у спортсменов высокой квалификации //
Теория и практика физ. Культуры. 1964. №10. С. 17-21.
Моисеев H.H. Алгоритмы развития. М.: Наука. 1987. 304 с.
Моисеев H.H. Экология человечества глазами математика. М.: Молодая
гвардия. 1988. 254 с.
Моисеев H.H., Александров В.В., Тарко A.M. Человек и биосфера. М.:
Наука. 1985. 271 с.
Морган Т.Г. Теория эволюции в современном освещении. М.-Л.: Госиздат.
1926.
Морган Т.Г. Экспериментальные основы эволюции. М.-Л.: Биомедгиз. 1936.
250с.
Мурадян Х.К. Распределение видовой продолжительности жизни в
различных таксономических группах животных // Журнал общ.
биологии. 1989. Т.50. №1. С.22-26.
Нагорный A.B., Никитин В.М., Буланкин H.H. Проблемы старения и
долголетия. М.: Медгиз. 1963. 755 с.
Назаров В.И. Финализм в современном эволюционном учении. М.: Наука.
1984. 284 с.
Наумов H.H. Зоология позвоночных. М.: Просвещение. 1965. 463 с.
Наумов Н.П., Карташев H.H. Зоология позвоночных. М.: Высшая школа.
1979.4.1.333с.
Несис К.Н. Первые ископаемые гребневики // Природа. 1984. №5. С. 115-116.
Несис К.Н. Кембрийская революция шла быстрее, чем думали // Природа.
1994. №12. С.94-95.
Никитенко М.Ф. Головной мозг, эволюция и систематика млекопитающих //
Внутривидовая изменчивость наземных прозвоночных животных и
микроэволюция / ред. С.С.Шварц. Свердловск: Изд. АН СССР. 1965.
С.119-136.
Никитенко Μ.Φ. Эволюция и мозг. Минск: Наука и техника. 1969. 342 с.
Николаев А.Я. О термодинамике живых систем // Биофизика. 1966. Т.11.№3.
С.551-554.
Николаев Л.А. Основы физической химии биологических процессов. М.:
Высшая школа. 1976. 260 с.
Николис Г., Пригожин И.Р. Самоорганизация в неравновесных системах. М.:
Мир. 1979.512с.
Новосельцев В.Н. Теория управления и биосистемы. М.: Наука. 1978. 320 с.
Обручев Д.В. (Ред.) Основы палеонтологии. Справочник для палеонтологов
и геологов СССР. Т.Н. Бесчелюстные и рыбы М.: Наука. 1964. 521 с.
Озернюк Н.Д. Энергетический обмен в раннем онтогенезе рыб. М.: Наука.
1985. 175 с.
Озернюк Н.Д. Принцип энергетического минимума в онтогенезе и
устойчивость процессов развития // Журнал общей биологии. 1988.
Т.49. №4. С.552-562.
Озернюк Н.Д. Принцип минимума энергии в онтогенезе и
канализированность процессов развития // Онтогенез. 1989. Т.20.№2.
С. 117-127.
Озернюк Н.Д. Механизмы адаптации. М.: Наука. 1992. 272 с.
Озернюк Н.Д., Булгакова Ю.В. Стандартный метаболизм у рыб и
круглоротых: эволюционные и экологические закономерности //
Известия РАН. Сер.биол. 1997. №5. С.571-579.
Озернюк Н.Д., Зотин А.И., Юровицкий Ю.Г. Оогенез как модель процесса
уклонения живой системы от стационарного состояния // Онтогенез.
1971. Т.2. №6. С.565-571.
Опарин А.И. Возникновение жизни на Земле. М.: Изд-во АН СССР. 1957.
195с.
Павлова Е.В. Дыхание планктонных животных Черного моря //
Биологические исследования Черного моря и его промысловых
ресурсов / Ред. В.А.Водяницкий. М.: Наука. 1968. С.71-75.
Павлова Е.В. Метаболизм средиземноморского зоопланктона //
Биологическая структура и продуктивность сообществ Средиземного
моря / Ред. В.Е.Заика. Киев: Наукова Думка. 1975. С. 124-144.
Паевский В.А. Демография птиц. Л.: Наука. 1985а. 285 с.
Паевский В.А. Масса тела и продолжительность жизни птиц // Вестник зоол.
19856. №2. С.70-72.
Пантелеев П.А. Биоэнергетика мелких млекопитающих. М.:Наука. 1983.271с.
Парамонов A.A. Пути и закономерности эволюционного процесса (основные
принципы филогенетики) // Современные проблемы эволюционной
теории / Ред. В.И.Полянский, Ю.И.Полянский. Л.: Наука.. 1967.
С.342-441.
Пасынский А.Г. Биофизическая химия. М.: Высшая школа. 1963. 432 с.
Перельман А.И. Геохимия биосферы. М.: Наука. 1973. 167 с.
Печуркин Н.С. Энергетические аспекты развития надорганизменных систем.
Новосибирск: Наука. 1982. 113 с.
Плавильщиков H.H. Определитель насекомых. М.: Гос. Учебно-педагогич.
Изд-во. 1957. 548 с.
Поплавский Р.П. Термодинамические модели информационных процессов //
Успехи физ. наук. 1975. Т. 115, №3. С.465-501.
Поплавский Р.П. Термодинамика информационных процессов. М.: Наука.
1981.255с.
Преснов Е.В. Потенциальный характер критерия эволюции термодинамики
необратимых процессов // Журнал физ. химии. 1973. Т.47, №11.
С.2902-2904.
Преснов Е.В. Формализм неравновесной феноменологической
термодинамики // Термодинамика биологических процессов / Ред.
А.И.Зотин. М.: Наука. 1976а. С.25-42.
Преснов Е.В. Усиленный критерий эволюции и биология развития //
Термодинамика биологических процессов / Ред. А.И.Зотин. М.:
Наука. 19766. С.73-76.
Преснов Е.В. Феноменологические уравнения и критерий эволюции
неравновесной термодинамики // Переходные процессы в
биологических системах (тезисы) / Ред. А.И.Зотин. Москва. 1977.
С.21-25.
Пригожин И. Проблема эволюции в термодинамике необратимых явлений //
Возникновение жизни на Земле / Ред. А.И.Опарин. М.: Изд-во АН
СССР. 1959. С.408-416.
Пригожин И. Введение в термодинамику необратимых процессов. М.: Изд-во иностр. литер. 1960. 127 с.
Пригожин И. От существующего к возникшему. Время и сложность в
физических науках. М.: Наука. 1983. 327 с.
Приц А. К. Принцип стационарных состояний открытых систем и динамика
популяций. Калининград: Изд-во гос. ун-та. 1974. 123 с.
Проссер Л. (Ред.) Сравнительная физиология животных. М.: Мир. 1977а. Т.1.
608с.
Проссер Л. (Ред.) Сравнительная физиология животных. М.: Мир. 19776.
Т.2. 571 с.
Путилов К.А. Термодинамика. М.: Наука. 1971. 375 с
Пчелинцев В.Ф., Коробков И.А.. (Ред.) Основы палеонтологии. Справочник
для палеонтологов и геологов СССР. Т.4. Моллюски - брюхоногие.
М.: Гос. научно-техн. изд-во литературы по геологии и охране недр.
1960. 360 с.
Расницын А.П. О несводимости макроэволюционных процессов к
микроэволюционным. // Философские проблемы эволюционной
теории/Ред. М.Е.Быховский и др. М.: Наука. 1971. 4.2. С. 171-178.
Расницын А.П. История палеоэнтомологии и история насекомых // Природа.
1990. №6. С.66-80
.Расе Т.С. Тип позвоночных. Общий очерк. // Жизнь животных / Ред.
Т.С.Расс. М.: Просвещение. 1971. Т.4. С.7-11
Решетников Ю.С., Котляр А.Н., Расе Т.С., Шатуновский М.И. Пятиязычный
словарь названий животных. Рыбы. М.: Русский язык. 1989. 734 с.
Рогозкин В.А., Афар Я., Машанский В.Ф. Ферментативная активность в
ультраструктуре митохондрий при гипертрофии мышц // Биохимия.
1964. Т.29. №5. С.905-909.
Родендорф Б. Б. (Ред.) Основы палеонтологии. Справочник для
палеонтологов и геологов СССР. Т.9. Членистоногие - трахейные и
хелицеровые. М.: Изд-во АН СССР. 1962. 560 с.
Рождественский А.К., Татаринов Л.П. (Ред.) Основы палеонтологии.
Справочник для палеонтологов и геологов СССР. Т. 12. Земноводные,
пресмыкающиеся, птицы. М.: Наука. 1964. 721 с.
Розен Р. Принцип оптимальности в биологии. М.: Мир. 1969. 215 с.
Росс Г., Росс Ч., Росс Д. Энтомология. М.: Мир. 1985. 676 с.
Роуз Д.Дж., Кларк М. Физика плазмы и управляемые термоядерные реакции.
М.: Госатомиздат. 1963. 487 с.
Рубин А.Б. Термодинамика биологических процессов. М.: Изд-во МГУ. 1984.
240с.
Руденко А.П. Теория саморазвития открытых каталитических систем. М.:
Изд-во МГУ. 1969. 276 с.
Руженцев В.Е. (Ред.) Основы палеонтологии. Справочник для палеонтологов
и геологов СССР. Т. 5. Моллюски - головоногие, I:
наутилоидеи,.бактритоидеи, аммоноидеи (агониатиты, гониатиты,
климении). М.: Изд-во АН СССР. 1962. 438 с.
Руттенбург С.О. Химическая терморегуляция у некоторых видов крыс в
связи с их географическим распространением // Опыт изучения
регуляций физиологических функций / Ред. К.М.Быков. М./Л.: Изд-во АН СССР. 1953. Т.2. С.35-45.
Рябушко В.И. Энергетический обмен иглокожих Баренцева и Японского
морей // Биология моря. 1978. №2. С.56-63.
Рябушко В.И. Энергетический обмен иглокожих. Автореферат докторской
дисс., Минск, 1992. 50 с.
Рябушко В.И., Пропп Л.Н. Интенсивность дыхания, азотного и фосфорного
метаболизма у иглокожих Южно-Китайского моря // Биология моря.
1985. №6. С.42-46.
Светницкий И.И. К обоснованию биоэнергетической целенаправленности
структур и функций живых систем. Деп. №3204. М.: ВИНИТИ, 1977.
Светницкий И.И. Биоэнергетическая направленность как общебиологическая
закономерность. Препринт. Пущине. 1980.
Светницкий И.И. Биоэнергетическая направленность и проблемы экологии.
Препринт. Пущине. 1981. 12 с.
Северцов А.Н. Главные направления эволюционного процесса. Прогресс,
регресс и адаптации. М.: Думнов. 1925. 83 с.
Северцов А.Н. Главные направления эволюционного процесса.
Морфобиологическая теория эволюции. М.-Л.: Биомедгиз. 1934. 150
с.
Северцов А.Н. Морфологические закономерности эволюции. М.-Л.: Изд-во
АН СССР. 1939.610с.
Северцов A.C. Основы теории эволюции. М.: Изд-во МГУ, 1987. 320 с.
Северцов A.C. Направленность эволюции. М.: Изд-во МГУ, 1990. 270 с.
Сегаль А.Н. Терморегуляция у нутрии (Myocastor coypus) летом // Зоол.
журнал. 1978. Т.57, №12. С. 1878-1883.
Сидоренко A.B. Новое в учении о биосфере. М.: Наука, 1984. 15 с.
Симпсон Дж.Г. Темпы и формы эволюции. М.: Изд-во иностр. литер. 1948.
369с.
Слоним А.Д. Животная теплота и ее регуляция в организме млекопитающих.
М./Л.: Изд-во АН СССР. 1952. 325 с.
Слоним А.Д. Основы общей экологической физиологии млекопитающих.
М./Л.: Изд-во АН СССР. 1961. 432 с.
Слоним А.Д. Термические адаптации и температурная компенсация у
гомойотермных организмов // Экологическая физиология животных /
Ред. А.Д.Слоним и др. Л.: Наука. 1982а. Ч.З. С.41-47.
Слоним А.Д. Адаптации к холоду и высоким широтам // Экологическая
физиология животных / Ред. А.Д.Слоним и др. Л.: Наука. 19826. Ч.З.
С.67-80.
Слоним А.Д. Стратегия температурных адаптации в разных климатических
зонах // Экологическая физиология животных / Ред. А.Д.Слоним и
др. Л.: Наука. 1982в. Ч.З. С. 188-190.
Слоним А.Д. Физиологические изменения в организме человека в горах //
Экологическая физиология животных / Ред. А.Д.Слоним и др. Л.:
Наука. 1982г. Ч.З. С.393-398.
Слоним А.Д. Эволюция терморегуляции. Л.: Наука. 1986. 76 с.
Смирнов П.К. Некоторые проявления внутривидовой эколого-
физиологической дифференциации у мышевидных грызунов //
Вестник ЛГУ. 1957. №21. Сер.биол. Вып.4. С. 106-114.
Соколов B.C. (Ред.) Основы палеонтологии. Справочник для палеонтологов
и геологов СССР. Т.2. Губки, археоциаты, кишечнополостные, черви.
М.: Изд-во АН СССР. 1962. 485 с.
Соколов B.C. Палеонтология и эволюция биосферы // Палеонтология и
эволюция биосферы / Ред.А.Н.Олейников и др. Л.: Наука. 1983. С.4-
17.
Соколов B.C. Биосфера: понятие, структура, эволюция // В.И.Вернадский и
современность / Ред. А.Г.Назаров. . М.: Наука. 1986. С.96-123.
Соколов В.Е. Систематика млекопитающих. М.: Высшая школа. 1973. Т.1.
430с.
Соколов В.Е. Систематика млекопитающих. М.: Высшая школа. 1977. Т.2.
494с.
Соколов В.Е. Систематика млекопитающих. М.: Высшая школа. 1979. Т.З.
528с.
Соколов В.Е. Словарь названий животных. Млекопитающие. М.: Русский
язык. 1984. 352 с.
Старобогатов Я.И. О соотношении между микро- и макроэволюции //
Дарвинизм: история и современность / Ред.Э.И.Колчинский,
Ю.И.Полянский. Л: Наука. 1988. С. 138-145.
Старобогатов Я.И., Левченко В.Ф. Эксцентрическая концепция
макроэволюции//Журнал общ. биологии. 1993. Т.54, №4. С.389-407.
Стеттнер Л., Матиньяк К. Мозг птиц // Птицы / Ред. Л.С.Степанян,
С.Н.Хаютин. М.: Мир. 1983. С.211-220.
Стрельников И.Д. О соотношении величины мозга и теплопродукции у
грызунов // Докл. АН СССР. 1953. Т.88, №2. С.377-380.
Стрельников И.Д. Анатомо-физиологические основы видообразования
позвоночных. Л.: Наука. 1970а. 367 с.
Стрельников И.Д. Закон взаимосвязи видообразования животных с
величиной их тела, мозга и энергией физиологических процессов //
Зоол. журнал. 19706. Т.49. №4. С.534-559.
Сутт Т. Проблема направленности органической эволюции. Таллин: Валгус.
1977. 140 с.
Суханова K.M. Температурные адаптации у простейших. Л.: Наука, 1968.
267с.
Сущеня Л.М. Интенсивность дыхания ракообразных. Киев: Наукова думка,
1972. 196 с.
Сырников Ю.П. О применимости второго начала термодинамики к живой
материи//Биофизика. 1965. Т. 10. №6. С. 1102-1103.
Татаринов Л.П. Очерки по теории эволюции. М.: Наука, 1987. 251с.
Тахтаджан А.Л. Пути приспособительной эволюции растений //
Ботанический журнал. 1951. Т.36, №3. С.231-239.
Тахтаджан А.Л. Происхождение и расселение цветковых растений. Л.:
Наука. 1970. С. 13-15.
Тахтаджан А.Л. Четыре царства органического мира // Природа. 1973. №2. С.
22-32.
Терентьев П.В. Герпетология. М.: Высшая школа. 1961. 336 с.
Тимирязев К.А. Чарльз Дарвин и его учение. Сочинения. М: Сельхозгиз.
1939. T.VII. 675 с.
Тимофеев-Ресовский Н.В. О взаимоотношениях микро- и макроэволюции //
Микро- и макроэволюция / Ред. К.Л.Паавер. Тарту: АН Эстонской
ССР. 1980. С.51-58.
Тимофеев-Ресовский Н.В., Воронцов H.H., Яблоков A.B. Краткий очерк
теории эволюции. М.: Наука. 1969. 408 с.
Тихонова Н.С., Дорошкевич В.И. Исследование газообмена у Limnodrilus
udekemianus в воздушной среде // Опыт эколого-физиологического
изучения загрязнения водной среды / ред. А.Д.Слоним и др. Фрунзе:
Илим. 1976. С.22-28.
Тринчер К.С. Биология и информация. Элементы биологической
термодинамики. М.: Наука. 1965. 119 с.
Тринчер К.С. По поводу рецензий и дискуссионных статей, касающихся
"Элементов биологической термодинамики" // Биофизика. 1966. Т. 11,
№6.0.1097-1100.
Тыщенко В.П. Основы физиологии насекомых. Часть 1. Л.: Изд-во ЛГУ. 1976
Умнов A.A. Применение статистических методов для оценки параметров
эмпирических уравнений, описывающих взаимосвязь между
энергетическим обменом и массой тела животного // Журнал общ.
биологии. 1976. Т.37. №1. С.71-86.
Уоддингтон К. Основные биологические концепции // На пути к
теоретической биологии / Ред. Б.Л.Астауров. М.: Мир. 1970. С. 11-38.
Уоллэсъ А. Дарвинизмъ. Изложенше теорш естественаго подбора и
нъкоторихъ изъ ея приложенш. М.: Сытин. 1898. 753 с.
Филатова З.А. Класс Двустворчатые (Bivalvia) // Жизнь животных.
Беспозвоночные / Ред.Л.А.Зенкевич. М.: Просвещение. Т.2. С.95-155.
Филипченко Ю.А. Эволюционная идея в биологии. М.: Наука. 1977. 227 с.
Фокс Р. Энергия и эволюция жизни на Земле. М.: Мир. 1992. 216 с.
Фокс С., Дозе Л. Молекулярная эволюция и возникновение жизни. М.: Мир.
1975. 374 с.
Хаазе Р. Термодинамика необратимых процессов. М.: Мир. 1967. 544 с.
Хазен A.M. Особенности применения второго начала термодинамики к
описанию работы мозга // Биофизика. 1991. Т.36. №4. С.714-724.
Хазен А. М Происхождение и эволюция жизни и разума с точки зрения
синтеза информации//Биофизика. 1992. Т.37. №1. С. 105-122.
Хазен A.M. Принцип максимума производства энтропии и движущая сила
прогрессивной биологической эволюции // Биофизика. 1993а. Т.38,
№3. С. 531-551.
Хазен A.M. Почему самопроизвольно возникает и прочитывается
генетическая информация // Теоретическая биология. 19936. Вып.5.
48с.
Хазен A.M. Введение меры информации в аксиоматическую базу механики.
Москва. 1996. 48 с.
Ханин М.А., Дорфман Н.Л., Бухаров И.Б., Левадный В.Г. Экстремальные
принципы в биологии. М.: Наука. 1978. 256 с.
Хаскин В. В. Энергетика теплообразования и адаптация к холоду.
Новосибирск: Наука. 1975. 200 с.
Хаскин В.В. Энергетический обмен // Экологическая физиология животных /
Ред. А.Д.Слоним и др. Л.: Наука. 1981. 4.2. С.379-407.
Хаскин В.В. Биохимические механизмы адаптации к холоду // Физиология
терморегуляции / Ред. К.П.Иванов и др. Л.: Наука. 1984. С.237-266.
Хлебович Т.В. Интенсивность дыхания у инфузорий разного размера //
Цитология. 1974. Т. 16. №1. С. 103-106.
Хлебович Е.В., Винберг Г.Г. Уровень энергетического обмена у
многоклеточных беспозвоночных животных и у простейших //
Доклады АН СССР, 1984. Т.274. №2. С.497-499.
Холодковский H.A. Учебник зоологии. Петроград: Девриен. 1918. 546 с.
Холодковский H.A. Курс энтомологии теоретической и прикладной. М.-Л.:
Сельхозгиз. 1929. 400 с.
Цейтлин В.Б. Энергетика глубоководных пелагических сообществ. М.:
Наука. 1986. 113с.
Циглер Г. Экстремальные принципы термодинамики необратимых
процессов и механики сплошной среды. М.: Мир. 1966. 135 с.
Чернавский Д.С. Термодинамика и биология // Теоретическая и
экспериментальная биофизика / Ред.А.К.Приц. Калининград. 1975.
Т.5. С.3-27.
Чернышева Н.Е. (Ред.) Основы палеонтологии. Справочник для
палеонтологов и геологов СССР. Т. 8. Членистоногие
трилобитообразные и ракообразные. М.: Гос. научно-техн. изд-во
литературы по геологии и охране недр. 1960. 515 с.
Шамбадаль П. Развитие и приложение понятия энтропии. М.: Наука. 1967.
278с.
Шахпаронов М.И. Неравновесная термодинамика и принцип наименьшего
действия // Термодинамика необратимых процессов / Ред.
А.И.Лопушанская. М.: Наука. 1987. С.87-96.
Шварц С.С. К методологии исследования макроэволюционного процесса //
Научи, труды ветер, академии. 1976. Т.84. С. 11-25.
Шварц С.С. Экологические закономерности эволюции. М.: Наука. 1980.278с.
Шилов H.A. Регуляция теплообмена у птиц. М.: Изд-во МГУ. 1968. 252 с.
Шилов H.A. Физиологическая экология животных. М.: Высшая школа. 1985.
328с.
Шилова Е.И. О понятии "биосфера", его значение и современное толкование
//Вестник ЛГУ. 1983. Биология. №3, Вып.1. С.5-16.
Шипунов Ф.Я. Организованность биосферы. М.: Наука. 1980. 291 с.
Шкловский И.С. Вселенная, жизнь, разум. М.: Наука. 1987. 320 с.
Шмальгаузен И.И. Пути и закономерности эволюционного процесса. М./Л.:
Изд-во АН СССР. 1939. 231 с.
Шмальгаузен И.И. Факторы эволюции. Теория стабилизирующего отбора.
М./Л.: Изд-во АН СССР. 1946. 396 с.
Шмальгаузен И.И. Происхождение наземных позвоночных. М.: Наука. 1964.
271с.
Шмальгаузен И.И. Проблемы дарвинизма. Л.: Наука. 1969. 493 с.
Шмидт-Ниелсен К. Физиология животных: Приспособление к среде. М.:
Мир. 1982. 414 с.
Шмидт-Ниелсен К. Размеры животных: Почему они так важны? М.: Мир.
1987. 159 с.
Шноль С.Э. Физико-химический аспект процесса биологической эволюции.
Возможная детерминированность основных этапов эволюции
физическими свойствами эволюционирующих систем // Журнал общ.
биологии. 1973. Т.34, №3. С.ЗЗ 1-347.
Шноль С.Э. Физико-химические факторы биологической эволюции. М.:
Наука. 1979. 262 с.
Шредингер Э. Что такое жизнь с точки зрения физики? М.: Мир. 1947. 128 с.
Шульман Г.Е., Аболмасова Г.И., Муравская З.А. Физико-биохимические
особенности эпипелагических кальмаров Индийского и
Атлантического океанов // Систематика и экология головоногих
моллюсков / Ред.Я.И.Скоробогатов, К.Н.Несис. Л.: Изд-во АН СССР.
1983.С.120-121.
Щербаков А.П. Дыхание и температурная адаптация у речной миноги
(Lampetra fluviatilis) II Бюллютень эксп. биол. и мед. 1937. Т.З. №3.
С.286-289.
Щукина М.Я., Дорошкевич В.И. Влияние температурной адаптации на
потребление кислорода Limnodrilus udekemianus // Опыт эколого-
физиологического изучения загрязнения водной среды / ред.
А.Д.Слоним и др. Фрунзе: Илим. 1976. С.29-34.
Эбелинг В. Образование структур при необратимых процессах: введение в
теорию диссипативных структур. М.: Мир. 1979. 279 с.
Эберзин А.Г. (Ред.) Основы палеонтологии. Справочник для палеонтологов
и геологов СССР. Т.З. Моллюски - панцирные, двустворчатые,
лопатоногие. М.: Изд-во АН СССР. 1960. 300 с.
Яблоков A.B. О разных формах прогрессивного развития в органической
природе // Проблемы эволюции Новосибирск: Наука. 1968. Т.1. С.98-
115.
Яблоков A.B. Существует ли специальный механизм макроэволюции? //
Микро- и макроэволюция / Ред. К.Л.Паавер. Тарту: АН Эстонской
ССР. 1980. С.62-65.
Яблоков A.B. От познания эволюции к управлению // Наука и жизнь. 1984.
№11,0.13-15.
Яблоков A.B., Белькович В.М., Борисов В.И. Киты и дельфины. М.: Наука.
1972. 472 с.
Яблоков A.B., Юсуфов А.Г. Эволюционное ученье. М.: Высшая школа. 1998.
331с.
Якименко М.А. Длительная адаптация организма человека и животных к
холоду // Физиология терморегуляции / Ред. К.П.Иванов и др. Л.:
Наука. 1984. С.223-236.
Яковенко В.А. Влияние горного климата на основной обмен человека //
Бюлл. НИИ курортологии и физиотерапии. 1938. №3/4. С. 1-2.
Яковлева К. К. Интенсивность дыхания у некоторых черноморских медуз //
Тр-ды Севастопольской биол. станции. 1964. Т. 17. С.364-367.
Ярославцева Л.М., Сергеева Э.П., Зотин А.И. Определение значений
температуры и солености, оптимальной для дробления яиц морского
ежа // Биология моря. 1992. №3/4. С.83-91.
Adams R.N. Energy and Structure - A Theory of Social Power. Austin: Univ.
Texas Press. 1975.
Agrell I. Some experiments concerning thermal adjustment and respiratory
metabolism in insects // Arkiv Zool. 1947. V.39A, N3. P. 1-48.
Alekseeva T.A., Zinichev V.V., Zotin A.I. Energy criteria of reliability and
stability of development // Acta Zool. Fennica. 1992. V.191. P.157-163.
Allen G.D. Quantitative studies on the rate of respiratory metabolism in Planaria.
II. The rate of oxygen consumption during starvation, feeding, growth, and
regeneration in relation to the method of susceptibility to potassium
cyanide as a measure of rate of metabolism // Amer. J. Physiol. 1919.
V.49. N3. P.420-473.
Allen M.D. Respiration rates of worker honeybees of different ages and at
different temperature // J. exp. Biol. 1959. V.36, N1. P.92-101.
Anderson J.F. Metabolic rates of spiders // Сотр. Biochem. Physiol. 1970. V. 33.
Nl.P.51-72.
Anderson J.F., Prestwich K.N. Respiratory gas exchange in spiders // Physiol.
Zool. 1982. V. 55. N1. P. 72-90.
Andrews R.M., Pough F.H. Metabolism of squamate reptiles: allometric and
ecological relationships // Physiol. Zool. 1985. V.58. N2. P.214-231.
Anraku M. Influence of the Cape Cod Canal on the hydrography and on the
copepods in burrards bay and cape cod bay, Massachusetts. 2. Respiration
and feeding // Limnol. Oceanogr. 1964. V.9, N2. P. 195-206.
Arai M.N. Oxygen consumption of fed and starved Aequorea victoria (Murbach
and Shearer, 1902) (Hydromedusae). // Physiol. Zool. 1986. V.59, N2.
P.188-193.
Arcos F.C., Sohal R.S., Sun Sh.-Ch., Argus M.F., Burch G.E. Changes in
ultrastructure and respiratory control in mitochondria of rat heart
hypertrophied by exercise // Exp. and Med. Path. 1968. V.8. N1. P.49-65.
Armstrong E.A Look at relative brain size in mammals // Neurosci. Lett. 1982.
V.34, N2. P.101-104.
Âstrand P.O. New records in human power // Nature. 1955. V.176. P.922-923.
Azzone G.F. Negentrpy and historical arrow of time. Thermodynamical and
informational aspects of Darwinian revolution // What is Controlling Life?
/ Eds. E.Gnaiger, F.N.Gellerich, V.Wyss. Innsbruck: Univ. Press. 1994.
P.38-44.
Bartholomew G.A., Burnhard M.C. Trachéal gases, respiratory gas exchange,
body temperature and flight in some tropical cicadas // J. exp. Biol. 1984.
V.lll.P.131-144.
Bartholomew G.A., Casey T.M. Body temperature and oxygen consumption
during rest and activity in relation to body size in some tropical beetles //
J. Thermal. Biol. 1977. V.2, N.4 P.173- 185.
Bartholomew G.A., Casey T.M. Oxygen consumption of moths during rest, pre-
flight warm-up, and flight in relation to body size and wing morphology //
J. exp. Biol. 1978. V.76. P. 11-25.
Bartholomew G.A., Lighten J.R.B., Feener D.H. Energetics of trail running, load
carriage, and emigration in the column-raiding army ant Eciton hamatum
II Physiol. Zool. 1988. V.61, N1. P.57-68.
Bartholomew G.A., Tucker V.A. Size, body temperature, thermal conductance,
oxygen consumption, and heart rate in australian varanid lizard // Physiol.
Zool. 1964. V.37, N4. P.341-354.
Bauer E. Lebensdauer, Assimilationsgrenze, Rubnersche Konstante und Evolution
// Biol. Z. 1931. Bd.51, H.l-2. S.74-80.
Baumgarten-Schumann D., Piiper J. Gas exchange in the gills of resting
unanaesthetized dogfish (Scyliorhinus stellaris} II Resp. Physiol. 1968.
V.5. N3.P.317-325.
Beamish F.W.H. Oxygen consumption of adult Petromyzon marinus in relation to
body weigt and temperature // J. Fish. Res. Bd Canad. 1973. V.30. N9.
P.1367-1370.
Beauregard Costa de O. Sur l'équivalence entre information et entropie dans la
rapport 1/k m2 // C. R. Acad. Sei. 1960. T.251, N25. P.2898-2900
Belman B.W., Giese A.C. Oxygen consumption of an asteroid and echinoid from
the Antarctic. Biol. Bull. 1974. V.146, N2. P.157-164.
Benedict F.G. Vital energetics: A study in comparative basal metabolism. Wash.:
Carnegie Inst., 1938. 215 p.
Bennett A.F., Dawson W.R. Metabolism // Biology of the Reptilia / Ed. Gans C.,
Dawson W.R. N.Y.:Acad. Press, 1976. V.5. P.127-223.
Bently PJ. Cutaneous respiration in the Congo eel Amphiuma means (Amphibia:
Urodela) // Сотр. Biochem. Physiol. 1975. V.50A. N1. P.121-124.
Bently P.J., Shield J.W. Respiration of some urodele and anuran Amphibia. II. In
air, role of the skin and lungs // Сотр. Biochem. Physiol. 1973. V.46A.
N1, P.29-38.
Beraldo M.J.A.H., Mendes E.G. The respiratory metabolism of the castes of two
leaf cutting ants, Atta laevigata (F.Smith, 1858) and Atta sexdens
rubropilosa (Forel,1908) // Сотр. Biochem. Physiol. 1981. V.68A, N2.
P.241-248.
Biot M.A. Variational Principles in Heat Transfer. Oxford: Clarendon. 1970. 185
P-Blagoveschenski A. On the relatin between biochemical properties and the degrees
of evolutionary development of jrganisms // Biol. gen. 1929. Bd.5. S.427-
438.
Biasing I. Experimentelle Untersuchungen ber den Umfang der ökologischen und
physiologischen Toleranz von Planaria alpina Dana und Planaria
gonocephala Duges //Zool. J. 1953. Bd.64, H.2. S.112-152.
Blem C.R., Ragan C.A., Scott L.S. The thermal physiology of two sympatric
treefrogs Hyla cinerea and Hyla chrysoscelis (Anura: Hylidae) // Сотр.
Biochem. Physiol. 1986. V.85A, N3. P.563-570.
Block W. Oxygen uptake by Nanorchestes antarcticus (Acari) // Oikos. 1976. V.
27. N 2. P. 320.
Block W. Oxygen consumption of the terrestrial mite Alaskozetes antarcticus
(Acari: Crypotostagmata) // J. exp. Biol. 1977. V.68. P.69-87.
Block W., Tilbrook PJ. Respiration studies on the Antarctic collembolan
Cryptopygus antarcticus II Oikos. 1975. V.26. N1. P. 15-25.
Blueweiss L., Fox H., Kudzma F., Nakachima D., Peters R., Sams S. Relationships
between body size and some life history parameters // Oecologia. 1978.
V.37, N2. P.257-272.
Bonin G. von. Brain weight and body weight in mammals // J. gen. Physiol. 1937.
V.16. N2. P.379-389.
Boolootian R.A., Cantor V.H A preliminary report on respiration, nutrition and
behavior of Arbacia punctulata II Life Sei. 1965. V.4 N16. P.1567-1571.
Borer K.T., Lane C.E. Oxygen requirement of Octopus briareus Robson at
different temperatures and oxygen concentration // J. exp. Mar. Biol. Ecol.
1971. V.7. N3. P.263-269.
Bouder-Rodoni R., Mangold K. Respiration and nitrogen excretion by the squid
Loligoforbest//Mar.Biol. 1989. V.103. N3. P.333-338.
Brafield A.E. The oxygen consumption of an echiuroid, Bonellia viridis Rolando //
J. exp. Biol. 1968. V.48. N2. P.427-434.
Brafield A.E., Chapman G. The oxygen consumption of Pennatula rubra Ellis and
some other anthozoans // Z. vergl. Physiol. 1965. Bd.50. N4. S.363-370.
Brand T. von. Biochemistry and Physiology of Endoparasites. Amsterdam:
Elsevier. 1979. 447 p.
Brand T. von, Bowman I.B.R. Studies on the aerobic and anaerobic metabolism of
larval and adult Taenia taeniaeformis II Exp. Parasit. 1961. V.U. N2. P.
276-297.
Brechme T., Gyorgy P. Untersuchungen über Höhenklimawirkung // Biochem.
Zischr. 1927. Bd.186. H.l/4. S.213-221.
Brett J.R., Sutherland D.B. Respiratory metabolism of pumpkinseed (Lepomis
gibbosus} in relation to swimming speed // J. Fish. Res. Bd. Canada. 1965.
V.22. N . P.405-409.
Briedis D., Seagrave R.C. Energy transformation and entropy production in living
systems. I. Application to embryonic growth // J. theoret. Biol. 1984.
V.110. N2. P.173-193.
Brillouin L. Life, thermodynamics, and cybernetics // Amer.Scientist. 1949. V.37.
4. P.554-568.
Brillouin L. Physical entropy and information // J. Appl. Phys. 1951. V.22 N4.
P.338-343.
Brody S. Bioenergetics and Growth. N.Y.: Reinhold. 1945. 1023 p.
Brody S., Proctor R.C. Growth and development, with special reference to
domestic animals. XXIII. Relation between basal metabolism and mature
body weight in different species of mammals and birds // Meissouri Univ,
agr. exp. St. Res. Bull. 1932. N166. P.89-101.
Brooks D.R. What's going on in evolution? A brief guide to some new ideas in
evolutionary theory // Canad. J. Zool. 1984. V.61. P.2637-2645.
Brooks D.R. Nonequilibrium thermodynamics and different axioms of evolution //
Acta Biotheor. 1986. V.35. N1/2. P.77-106.
Brooks D.R., Le Blond P.H., Gumming D.D. Information and entropy in a simple
evolution model // J. theoret. Biol. 1984. V.109. N . P.77-93.
Brooks D.R., Wiley E.O. Evolution as an entropie phenomenon // Evolutionary
theory: Paths and future / Ed. J.W.Pollard. N.Y.: Wiley. 1984. P.141-171.
Brooks D.R., Wiley E.O. Evolution as Entropy: Toward a Unified Theory of
Biology. Chicago: Chicago Univ. Press. 1988. 415 p.
Brown G.W. The metabolism of Amphibia // Physiology of the Amphibia / Ed.
J.A.Moore. N.Y.: Acad. Press. 1964. P. 1-98.
Brown W.L, Schick J.M. Bimodal gas exchange and the regulation of oxygen
uptake in Holothurians // Biol. Bull. 1979.V.156. N3. P.272-288.
Buchel W. Entropy and information in the Universe //Nature. 1967. V.158. P.319-
320.
Bucher T.L., Ryan M.J., Bartholomew G.A. Oxygen consumption during resting,
calling, and nest building in the frog Physalaemus pustulosus II Physiol.
Zool. 1982. V.55. N1. P.10-22.
Bullock Т.Н. Compensation for temperature in the metabolism and activity of
poikilotherms // Biol. Rev. 1955. V.30. N3. P.311-342.
Burrus S.K., Dill D.B., Burk D.L., Freeland D.V., Adams W.C. Observation at see
level and altitude of basal metabolic rate and related cardiopulmonary
function // Hum. Biol. 1974. V.46.N4. P.677-692.
Buskirk E.R., Taylor H.L. Maximal oxygen intake and its relation to body
composition, with special reference to chronic physical activity and
obesity // J. appl. Physiol. 1957. V.U. N1. P.72-78.
Butler P.J., Taylor E.W. The effect of progressive hypoxia on respiration in the
dogfish (Scyliorhinus canicula} at different seasonal temperatures // J. exp.
Biol. 1975. V.63. Nl.P.l 17-130.
Cahill K., Lustick S. Oxygen consumption and thermorégulation in Apis mellifera
workers and drones // Сотр. Biochem. Physiol. 1976. V.55 A. N4. P.355-
357.
Calder W. A. Consequences of body size for avian energetics // Avian Energetics /
Ed. R.A.Paynter. Cambridge.. 1974. P.86-98.
Calder W.A. Aging in vertebrates: allometric consideration of spleen size and
lifespan // Fed. Proc. 1976. V.35. N.2. P.96-97.
Calder W.A. Scaling of physiological processes in homeothermic animals // Ann.
Rev. Physiol. / Eds. I.S.Edelman, S.G.Schultz. Cflifornia: Palo Alto. 1981.
V.43. P.301-322.
Calder W.A. Body size, mortality, and longevity // J. theoret. Biol. 1983. V.102.
N2. P.135-144.
Calder W.A. Size, Function and Life History. Cambridge: Harvard Univ. 1984.
431 p.
Caldwell R.S. Thermal compensation of respiratory enzymes in tissues of the
goldfish (Carassius auratus) II Сотр. Biochem. Physiol. 1969. V.31. N1.
P.79-93.
Galloway N.O. Heat production and senescence // J. amer, geriatr. Soc. 1974.
V.22. N4. P.149-150.
Campbell B. Biological entropy pump // Nature. 1967. V.215. P. 1308.
Campbell C.M., Davies P.S. Thermal acclimation in the teleost, Blennius pholis
(L.) // Сотр. Biochem. Physiol. 1975. V.52A. N1. P.147-151.
Chan D.K.O., Wong T.M. Physiological adjustments to dilution of the external
medium in the lip-shark Hemiscyllium plagiosum (Bennett). 3. Oxygen
consumption and metabolic rates // J. exp. Zool. 1977. V.200. N1. P.97-
102.
Cannon W.B. Organization and physiological homeostasis // Physiol. Rev. 1929.
V.9. N3.P.399-431.
Cannon W.B. The Wisdom of the Body. L.: Kegan, Trench, Trubner. 1932. 312 p.
Carlisle D.B., Cloudsley-Thompson J.L. Respiratory function and thermal
acclimation in tropical invertebrates // Nature. 1968. V.218. P.684-685.
Cary S.C., Shank T., Stein J. Worms bask in extreme temperatures // Nature. 1998.
V.391.P.545-546.
Chappell M.A. Temperature regulation and energetics of the solitary bee Ctntris
pallida during foreging and intermale mate competition // Physiol. Zool.
1984. V.57. N2. P.215-225.
Chapman G. A note on the oxygen consumption of Renilla kolliker Pfeffer //
Сотр. Biochem. Physiol. 1972. V.42A. N4. P.863-866.
Chernova N.M., Byzova Ju.B., Chernova A.I. Relationship of number, biomass
and gaseous exchange rate indices in microarthropods in substrates with
various organic matter contents // Pedobiologia. 1971. V. 11. N 4. P.306-
314
Clark R.T., Otis A.B. Comparative studies on acclimatization of mice to carbon
monoxide and low oxygen // Amer. J. Physiol. 1952. V.I69. N2. P.285-
294.
Claridge P.N., Potter I.C. Oxygen consumption, ventilatory frequency and heart
rate of lampreys (Lampetra fluviatilis} during their spawning run // J. exp.
Biol. 1975. V.63. N1. P.193-206.
Clemens O.T. Ventilation and oxygen consumption in rosy finches and house
finches at sea level and high altitude // J. сотр. Physiol. 1988. V.158B.
P.5766
Cloudsley-Thompson J.L. Therrestrial invertebrates // Comparative Physiology of
Thermorégulation / Ed. G.C.Whittow. N.Y.: Acad. Press. 1970. V.l. P. 15-
77
Coenen-Stass D., Schaarschmidt D., Lamprecht I. Temperature distribution and
calorimetric determination of heat production in the nest of the wood and
Formica polyctena (Hymenjptera, Formicidae) // Ecology. 1980. V.61.
N.2. P.238-244.
Collins B.G. The ecological significance of thermoregulatory responses to heat
stress shown by two population of an Australian murid, Rattus fuscipes II
Сотр. Biochem. Physiol. 1973. V.44A. N4. P.I 129-1140.
Comfort A. The Biology of Senescence. New York: Elsevier. 1979. 414 p.
Cook S.F. The respiratory gas exchange in Termopsis nevadensis // Biol. Bull.
1932. V.63. N2. P.246-257.
Cope E.D. The Primary Factors of Organic Evolution. Chicago. 1896.
Cotter A.J.R. Re-investigation of size, axial gradient and light as factor affecting
the respiration of certain marine sponges // Сотр. Biochem. Physiol.
1978. V.60A. N2. P.I 17-122.
Courtney W.A.M., Newell R.C. Ciliary activity and oxygen uptake in
Branchiostoma lanceolatum (Pallas) // J. exp. Biol. 1965. V.43. N1. P.l-
12.
Cracraft J. A nonequilibrium theory for the rate-control of speciation and
extinction and origin of macroevolutionary patterns // Syst. Zool. 1982.
V.31.P.348-365.
Crile G., Quiring O.P. A record of the body weight and certain organ and gland
weight of 3690 animals // Ohio J. Sei. 1940. V.40. N5. P.219-259.
Culebras J.M., Fitzpatrick G.F., Brennan M.F., Bayden C.M., Moore F.D. Total
body water and exchangeable hydroen. II. A review of comparative data
from animals based on isotope dilution and desiccation with a report of
new data from the rat // Amer. J. Physiol. 1977. V.232. R60-R65.
Culter R.G. Evolutionary biology of senescence // The Biology of Aging / Eds.
J.A.Behnke, C.E.Finch, G.B.Moment. N.Y.: Plenum press. 1978. P.311-
360.
Culter R.G. Evolutionary biology of aging and longevity in mammalian species //
Aging and Cell Function. New-York: Plenum Press. 1984. P.l-147.
Daly J.J., Matthews H.M. Effect of weight and temperature upon oxygen
consumption of the land planarian Bipalium kewense // Physiol. Zool.
1982. V.55. N2. P.148-154.
Davies P.S. Physiological ecology of Patella. 1. Effect of body size and
temperature on metabolic rate // J. Mar. Biol. Assoc. UK. 1966. V.46. N3.
P.647-658.
Davies P.S. Physiological ecology of Patella. 2. Effect of environmental
acclimation on the metabolic rate // J. Mar. Biol. Assoc. UK. 1967. V.47.
Nl.P.61-74.
Davies P.S., Walkey M. The effect of body size and temperature upon oxygen
consumption of the cestode Schistocephalus solidus (Mller) // Сотр.
Biochem. Physiol. 1966. V.18. N2. P.415-425.
Dawson W.R., Bartholomew G.A. Relation of oxygen consumption to body
weight, temperature, and temperature acclimation in lizards U ta
stansburiana and Sceloporus occidentalis II Physiol. Zool. 1956. V.29.
Nl.P.40-51.
Dawson W.R., Hudson J.W. Birds // Comparative Physiology of Thermorégulation
/ Ed. G.C.Whittow. N.Y.: Acad. Press. 1970. V.l. P.224-310.
De Beer G.R. Embryos and Ancestors. Oxford. 1958.
De Donder Th. L'affinité. Paris: Gauthier, Villars, 1931. 88 p.
Dehnel P.A. Effect of temperature and salinity on the oxygen consumption of two
intertidal crabs // Biol. Bull. 1960. V.I 18. N2. P.215-249.
Dehnel P., Segal E. Acclimation of oxygen consumption to temperature in
American cockroach (Periplanete americana) II Biol. Bull. 1956. V.I 11.
Nl.P.53-64.
De Jong A.A. The influence of simulated solar radiation on the metabolic rate of
white-crowned sparrows // Condor. 1976. V.78. N2. P.174-179.
De Mont M.E., О Dor R.K. The effects of activity, temperature and mass on the
respiratory metabolism of the squid, Illex illecebrosus II J. mar. Biol.
Assoc. UK. 1984. V.64. N3. P.535-543.
Denbigh K.G. An Inventive Universe. London: Hutchinson. 1979. 250 p.
Dixon A.F.G. Metabolic acclimatization to seasonal changes in temperature in the
sycamore, Drepanosiphum platanoides (Sehr.), and lime aphid,
Eucallipterus tilliae L. // J. Oecologia. 1973. V.13. N3. P.205-210.
Dmi'el R., Borat A. Thermal behavior, heat exchange, and metabolism in the
desert snake Spalerosophis Clifford 11 Physiol. Zool. 1972. V.45. N1. P.78-
94.
Dresco-Derouet L. Etude d'Araignées et a'Opilions cavernicoles dans leur milieu.
I. Intensité respiratoire, premiers résultats // Annls Spéléol. 1969. V.24.
N3. P. 529-532.
Dresco-Derouet L. Le métabolisme respiratoire des scorpions. IV. Intensité
respiratoire de plusieurs espèces d'Afrique du Nord et des relations avec 1
'altitude // Bull. Soc. hist, natur. Toulouse. 1973. V.109. N3-4. P.235-240.
Dunn J.F. Low-temperature adaptation of oxidative energy production in cold-water fishes // Canad. J. Zool. 1988. V.66. N5. P.1098-1104.
Dye A.H., McCwynne L. The effect of temperature and season on the respiratory
rate of three psammolittoral gastropods // Сотр. Biochem. Physiol. 1980.
V.66A. Nl.P.107-111.
Economes A.C. Taxonomic differences in the mammalian life span - body weight
relationship and the problem of brain weight // Gerontology. 1980. V.26.
Nl.P.90-98.
Edgar W.D. Aspects of the ecological energetics of the wolf spider Pardosa
(Lycosa) lugubris (Walckenaer) // Oecologia. 1971. V. 7. N 2. P. 136-
.154.
Edney E.B. Acclimation to temperature in terrestrial isopods. 2. Heart rate and
standard metabolic rate // Physiol. Zool. 1964. V.37. N4. P.378-394.
Edney E.B. Water Balance in Land Arthropods / / Zoophysiology and Ecology.
Berlin: Springer. 1977. V.9. 282 p.
Edwards C.A., Irving L. The influence of temperature and season upon oxygen
consumption of the sand crab, Emerita talpoida Say // J. cell. сотр.
Physiol. 1943. V.21. N2. P. 169-182.
Egginton S., Sidell B.D. Changes in mitochondrial spacing in fish skeletal muscle
induced by environmtntal temperature // J. Physiol. 1986. V.373. N1.
P.78-88.
Eimer T. On Orthogenesis and the impotence of natural selection in species-formation. Part II. London. 1898.
Ellington W.R. Metabolic responses of the sea anemone Bunodosoma cavernata
(Bosc) to deelining oxygen tensions and anoxia // Physiol. Rev. 1982.
V.55. N3. P.240-249.
Elzen G.W. Oxygen consumption and water loss in the imported fire ant
Solenopsis invicta Buren // Сотр. Biochem. Physiol. 1986. V.84A. N1.
P.13-17.
Eppler R. Energetische Gesetze. Oekoenenergetische Betrachtungen von Urknall
bis zum Jahr 2000. Basel: Helbing, Lichtenhahn.1989.
Eyring H., Boyce R.P., Spikes J.D. Thermodynamics of living systems //
Comparative Biochemistry / Eds. M.Florkin, H.S.Mason. N.Y.: Acad.
Press. 1960. V.l. P. 15-73.
Famme P. Effect of shell valve closure by the mussel Mytilus edulis L. On the rate
of oxygen consumption in declining oxygen tension // Сотр. Biochem.
Physiol. 1980. V.67A. N1. P.167-170.
Farmanfarmaian A. The respiratory physiology of Echinoderms // Physiology of
Echinodermata / Ed. R.A.Boolootian. N.Y.: Wiley. 1966. P.245-265.
Feder M.E. Lunglessness, body size, and metabolic rate in salamanders // Physiol.
Zool. 1976. V.49. N4. P.398-406.
Feder M.E., Feder J.H. Diel variation in oxygen consumption in three dpecies of
Philippine gekkonid lizards // Copeia. 1981. P.204-209.
Fildes P. Some medical and other aspects of bacterial chemistry // Proc. Roy. Soc.
Med. 1934. V.28. N1. P.79-90.
Fish J.D., Preece O.S. The ecophysiological complex of Bathyporeia pilosa and B.
pelagica (Crustacea: Amphipoda). 1. Respiration // Mar. Biol. 1970. V.5.
Nl.P.22-28.
Fisher T.R. Oxygen uptake of the solitary tunicata Styela plicata II Biol. Bull.
1976. V.151.N2. P.297-305.
Fitzpatrick L.C., Atebara M.Y. Effects of acclimation to seasonal temperatures on
energy metabolism in the toad Bufo woodhousei II Physiol. Zool. 1974.
V.47. N2. P.I 19-129.
Fitzpatrick L.C., Brown A.V. Metabolic compensation to temperature in the
salamander Desmognathus ochrophaeus from a high elevation population
// Сотр. Biochem. Physiol. 1975. V.50A. N4. P.733-737.
Flower S.S. Contributions to our knowledge of the duration of life in vertebrate
animals. V. Mammals // Proc. Zool. Soc. London. 1931. Part I. P. 145-234.
Franz V. Die biologische Fortschritt. Die Theorie der organismengefchichtlichen
Vervollkommnung. Jena: Fischer. 1935. 82 S.
Franz V. Zum jetzigen Stand die Theorie vom biotechnischen Fortschritt in der
Pflanzen- und Tiergeschichte // Biol. Gener. 1951. Bd.19. H.3. S.368-385.
Freeman R.F.H. Volumetrie respirometer measurements of the oxygen
consumption of themdigenetic trematode Proctoeces subtenuis // Сотр.
Biochem. Physiol. 1962. V.7. N3. P. 199-209.
Freyberg M. Der Grundumsatz in mittleren Hohen // Schweiz, med. Woch. Sehr.
1956. Bd.86. H.21. S.620-636.
Friedenthal H. Ueber die Giltigkeit des Massenwirkung für den Energienmsatz der
lebendigen Substanz // Z. Physiol. 1910. Bd.24. H.8. S.321-327.
Galvao P.E., Tarasantchi J., Guertzenstein P. Heat production of tropical snakes in
relation to body weight and body surface // Amer. Jour. Physiol. 1965.
V.209. N3. P.501-506.
Garrod С. Variational principle for nonlinear steady flow // J. Non-Equilib.
Thermodyn. 1984. V.9. N2. P.97-106.
Gavrilov V.M., Dolnik V.R. Basal metabolic rate, thermorégulation and existence
energy in birds: world data // Acta 18th Congr. Intern. Ornithol. / Eds.
V.D.Ilychov, V.M.Gavrilov. M.: Nauka. 1985. P.421-466.
Gelineo S. Influence de milieu thermique d'adaptation sur la thermognese des
homothermes // Ann. Physiol. Physicochim. Biol. 1934a. T. 10. N5.
P.1082-1115.
Gelineo S. Influence de milieu thermique d'adaptation sur la courbe de la
thermorégulation // C.r. Soc. Biol. 1934b. T.40. P.l 17-129.
Gerald J.W., Cech JJ. Respiratory responses of juvenile catfish (Ictalurus
punctatus) to hypoxic conditions // Physiol. Zool. 1970. V.43. N1. P.47-
54.
Giese A.C., Farmanfarmain A., Hilden S., Doezema P. Respiration during the
reproductive cycle in the sea urchin, Strongylocentrotus purpuratus II
Biol. Bull. 1966. V.130. N2. P. 192-201.
Gladfelter W.B. General ecology of the cassiduloid urchin Cassidulus
caribbearum//Mar. Biol 1978. V.47. N2. P.149-160.
Glaser R. Einfhrung in die Biophysik. Jena: Fischer. 1976. 349 S. Goil M.M.
Physiological studies on trematodes - Fasciola gigantica rate of oxygen
consumption//Z. Parasit. 1961. Bd.20. H.6. S.568-571.
Golightly R.T., Ohmart R.D. Metabolism and body temperature of two desert
canids: coyotes and kit foxes // J. Mammal. 1983. V.64. N4. P.624-635.
Golley F.B., Gentry J.B. Bioenergetics of the southern harvester ant
Pogonomyrmex badius // Ecology. 1964. V.45. N.2. P.217-225.
Gollnick P.D., King D.W. Effect of exercise and training on mitochondria of rat
skeletal muscle // Amer. J. Physiol. 1969. V.216. N6. P. 1502-1509.
Gordon E.E., Spiegel M., Villee C.A. An inplin effect on sponge metabolism // J.
cell. сотр. Physiol. 1955. V.45. N3, P.479-483.
Gourbault N. Etude compare du mtabolisme respiratoire chez des Planaires piges
et hypoges II Сотр. Rend. hebd. Sanc. Sci.Paris. 1968. T.266D. N2.
P.145-148
Greenstone M.H., Bennett A.F. Foraging strategy and metabolic rate in spiders //
Ecology. 1980. V. 61. N 5. P. 1255-1259.
Griffiths RJ. Temperature acclimation in Actinia equina L. (Anthozoa) // J. exp.
Mar. Biol. Ecol 1977. V.28. N3. P.285-299.
Hadley N.F., Quinlan M.C., Kennedy M.L. Evaporative cooling in the desert
cicada: thermal efficiency and water/metabolic cost // J. exp. Biol. 1991.
V.159. P.269-283.
Hailey A., Davies P.M.C. Lifestyle, latitude and activity metabolism of natricine
snakes // J. Zool. 1986. V.209A. N . P.461-476.
Hails CJ. The metabolic rate of tropical birds // Condor. 1983. V.85. N1. P.61-65.
Halliday T.R., Adler K. (Eds.) The Encyclopedia of Reptiles and Amphibians.
New York: Equinox Book. 1986. 424 p.
Harris W.V. Termites. Their recognition and control. L.: Longman. 1971.
Hart J.S. Energy metabolism of the white-footed mouse, Peromyscus leucopus
noveboracensis, after acclimation at various environmental temperatures //
Canad. J. Zool. 1953. V.31. N2. P.99-105.
Hart J.S., Berger M. Energetics, water economy and temperature regulation during
flight // Proc.lSth Intern. Ornitol. Congr. Leiden: Brill. 1972. P. 189-199.
Harvey P.H. Allometric analysis and brain size // Intelligence and Evolutionary
Biology/Eds. HJJerison, IJerison. Berlin: Springer. 1988. P.199-210.
Harvey P.H., Bennett P.M. Brain size, energetics, ecology and history patterns //
Nature. 1983. V.306. P.314-315.
Heinrich B. Temperature regulation of the sphinx moth, Manduca sexta. I. Flight
energetics and body temperature during free and tethered flight // J. exp.
Biol. 1971. V.54. N1. P.141-152.
Heinrich B. Thermorégulation in bumblebees. II. Energetics of warm-up and free
flight//J. Сотр. Physiol. 1975. V.96. N.I. P.155-166.
Heinrich B. The mechanisms and energetics of honeybee swarm temperature
regulation // J. exp. Biol. 1981a. V.91. P.25-55.
Heinrich B. Insect Thermorégulation. New York: Wiley. 1981b.
Hemmingsen A.H. Energy metabolism as related to body size and respiratory
surface, and its evolution // Rep. Steno Memor. Hospital Nordisk
Insulinlabor. 1960. V.9. N2. P.7-110.
Hill B.J., Potter I.C. Oxygen consumption in ammocoetes of the lamprey
Ichthyomyzon hubbsi Raney // J. exp. Biol. 1970. V.53. N1. P.47-57/
Hock R.J., Roberts J.C. Effect of altitude on oxygen consumption of deer mice:
Relation of temperature and season // Canad. J. Zool. 1966. V.44. N3.
P.365-376.
Hofman M.A. Encephalization in mammals in relation to the size of cerebral
cortex // Brain Behav. Evol. 1982. V.20. N1-2. P.84-96.
Hofman M.A. Energy metabolism, brain size and longevity in mammals // Quart.
Rev. Biol. 1983. V.58. N4. P.495-512.
Holldobler В., Wilson E.O. The Ants. Berlin: Springer. 1990. Holloszy J.O.
Biochemical adaptation in muscle: Effects of exercise on mitochondrial
oxygen uptake and respiratory enzyme activity in skeletal muscle // J.
Biol. Chem. 1967. V.242. N9. P.2278-2282.
Hopcroft R.R., Ward B.D., Roff J.C. The relative significance of body surface and
cloacal respiration in Psolus fabricu (Holoturoidea: Dendrochirotida) //
Canad. J. Zool. 1985. V.63. N12. P.2878-2881.
Hopson J.A. Relative brain size and behaviour in archosaurian reptiles // Ann.
Rev. Ecol. System. / Eds. R.F.Johnston, P.W.Frank, C.B.Michener. 1977.
V.8. P.429-448.
Houlihan D.F., Duthie G. Measurement of oxygen consumption and sampling of
body fluids of echinoderms in situ II J. exp. mar. Biol. Ecol. 1981. V.51.
Nl.P.97-106.
Houlihan D.F., Innés A.J., Wells V.J., Wells J. Oxygen consumption and blood
gases of Octopus vulgaris in hypoxic condition // J. Сотр. Physiol. 1982.
V.148D. Nl.P.35-40.
Hughes G.M. On the respiration of Torpedo marmorata II J. exp. Biol. 1978.
V.73. P.85-105
Hughes G.M., Gaymer R., Moore M., Woakes J. Respiratory exchange and body
size in the Aldabra giant tortoise II J. exp. Biol. 1971. V.55. N3. P.651-
665.
Hughes G.M., Umezawa S.I. Oxygen consumption and gill water flow in the
dogfish Scyliorhinus canicula L. // J. exp. Biol. 1968. V.49. N3. P.557-
564.
Huxley J. Evolution. The Modern Synthesis. London: Allen.Unwin, 1944. 645 p.
Huxley J. The three types of evolutionary process // Nature. 1957. V.I80. P.454-
455.
Huxley J.S. The evolutionary process // Evolution as a Process / Eds. J.Huxley,
A.C.Hardy, E.B.Ford. London: Allen, Unwin. 1958. P. 1-23.
Hyman L.H. Physiological studies on Planaria. I. Oxygen consumption in relation
to feeding and starvation // Amer. J. Physiol. 1919a. V.49. N3. D.377-402.
Hyman L.H. Physiological studies on Planaria. I. Oxygen consumption in relation
to age (size) differences // Biol. Bull. 1919b. V.37. N6. P.388-403.
Hyman L.H. Physiological studies on Planaria. IV. A further study of oxygen
consumption during starvation // Amer. J. Physiol. 1920. V.53. N3. P.399-
420.
Ikeda T. Relationship between respiration rate and body size in marine plankton
animals as a function of the temperature of habitat // Bull. Fac. Fish.
Hokkaido Univ. 1970. V.21. N2. P.91-112.
Irving L., Krog H., Monson M. The metabolism of some Alaskan animals in
winter and summer // Physiol. Zool. 1955. V.28.N3. P.173-185.
Jaffé R., Fonok C. Energetics of social phenomena: physic applied to evolutionary
biology // Π Nuova Cimento. 1994. V.16D. N6. P.543-553.
Jankowsky H.D., Korn H. Der Einfluß der Adaptations temperature auf den
Mitochndrien gehalt des Fischmuskuls // Naturwissenschaften. 1965.
Bd.52. H.23. S.642-643.
Jenkins M.M. Respiration rates in Planarians. I. The use of the Warburg
respirometer in determining oxygen consumption // Proc. Oklahoma Acad.
Sei. 1960. V.40. P.35-40.
Jensen T.F. Annual production and respiration in ant populations // Oikos. 1978.
V.31.N2. P.207-213.
Jerison H.J. Quantitative analysis of evolution of the brain in mammals // Science,
1961. V.133.P.1012-1014.
Jerison H.J. Gross brain indices and the analysis of fossil endocasts // Advances in
Primatology. V.I. Primate Brain / Eds. C.R.Noback, W.Montagna. New
York: Appleton-Century-Crofts. 1970. P.225-244.
Jerison H.J. Evolution of the Brain and Intelligence. New York: Acad. Press.
1973.482p.
Jerison H.J. Evolutionary biology of intelligency: the nature of the problem //
Intelligence and Evolutionary Biology / Eds. H.J.Jerison, I.Jerison. Berlin:
Springer. 1988. P.l-11.
Jerison HJ., Jerison I. (Eds.) Intelligence and Evolutionary Biology. Berlin:
Springer. 1988. 481 p.
Job S.V. The respiratory metabolism of Tilapia mossambica. II. The effect of size,
temperature, salinity and partial pressure of oxygen // Mar. Biol. 1969.
V.3. N . P.222-226
Johansen K. Cardie output in the large Ctphalopod, Octopus dofleini II J. exp.
Biol. 1965. V.42. N3. P.475-480.
Johansen K., Lenfant C. Gas exchange in the cephalopod, Octopus dofleini II
Amer. J. Physiol. 1966. V.210. N4. P.910-918.
Johansen K., Lenfant C., Hanson D. Gas exchange in the lamprey, Entosphenus
tridentatus II Сотр. Biochem. Physiol. 1973. V.44A. N1. P.107-119.
Johansen K., Petersen J.A. Gas exchange and active ventilation in a starfish,
Pteraster tasselatus IIZ. vergl. Physiol. 1971. Bd.71, H.4. S.365-381
Johansen K., Redmond J.R., Bourne G.B. Respiratory exchange and transport of
oxygen in Nautilus pompilus II J. exp. Zool. 1978. V.205. N1. P.27-36.
John T.L. Respiratory metabolism // Protozoa in Biological Research London:
Columbia Univ. 1941. P.352-403.
Johnston I.A., Maitland B. Temperature acclimation in crucian carp Carassius
carassius L., morphometric analysis of muscle fibre ultrastructure // J.
Fish. Biol. 1980. V.17. N1. P.I 13-125.
Jones M.L. Longevity of captive mammals // Zool. Gart. 1982. V.52. N2. P. 113-
128.
Jongeling T.B. On an axiomatization of evolutionary theory // J. theoret. Biol.
1985. V.I 17. N4. P.529-543.
Jürgens K.D., Prothero J.W. Scaling of mximal lifespan in bats // Сотр. Biochem.
Physiol. 1987. V.88A. N2. P.361-367.
Jürgens K.D., Prothero J. Life time energy budgets in mammals and birds // Сотр.
Biochem. Physiol. 1991. V.100A. N3. P.703-709.
Kammer A.E., Heinrich B. Metabolie rates related to muscle activity in
bumblebees // J. exp. Biol. 1974. V.61. N1. P.219-227.
Kale R.D., Rao K.P. Studies on neurohumoral induction of compensatory
mechanisms in thermal acclimation of poikilotherms. 1. Effect of injection
of CNS extract on the metabolism of two species of earthworms // J. exp.
Biol. 1973. V.59. N3. P.655-664.
Katchalsky A., Curran P.P. Nonequilibrium Thermodynamics in Biophysics.
Cambridge (Mass.): Harvard Univ.press, 1965. 248 p.
Keister M., Buck J. Respiration: some exogenous and endogenous effects on rate
of respiration // The Physiology of Insects / Ed. M.Rockstein. N.Y.:
Acad.Press. 1974. V.VI. P.469-509.
Kendeigh S.C., Dol'nik V.R., Gavrilov V.M. Avian energetics // Carnivorous birds
in ecosystems. Intern, biol. program. Cambridge: Univ.Press, 1977. V.12.
P. 127-204.
Kincannon E.A. The relations between body weight and habitat temperature and
the respiratory rate of Tonicella lineats (Wood, 1815) (Vjllusca:
Polyplacophora) // Veliger. 1975. V.18 (Suppl.). P.87-93.
King J.K., Earner D.S. Energy metabolism, thermorégulation and body
temperature // Biology and comparative physiology of birds / Ed.
A.J.Marshall. N.Y.: Acad. Press. 1961. V.2. P.215-288.
Kirby M., Kirby E. The Sea and its Wonders. London: Nelson and Sons.
1907.304p.
Kleiber M. Body size and metabolic rate // Physiol. Rev. 1947. V.27. N3. P.511-
541.
Kleiber M. The Fire of Life. An Introduction to Animal Energetics. N.Y.: Wiley.
1961.454p.
Knight A.W., Caufin A.R. Oxygen consumption of several species of stoneflies
(Plecoptera) // J. Insect Physiol. 1966. V.12. N3. P.347-355.
Knight B.C.J.G. Bacterial nutrition // Med. Res. Council (Brit.), Spec. Rep. Ser.
1936. N210. P.l-182.
Krogh A. The Respiratory Exchange of Animals and Man. London: Longman,
Green. 1916. 173 p.
Krüger F. Stoffwechsel und Wachstum bei Scyphomedusen // Helhol. wiss.
Meeresunters. 1968. Bd.18. N4. P.367-383.
Krüger K., Prinzinger R.,Schuchmann Torpor and metabolism in hummingbirds //
Сотр. Biochem. Physiol. 1982. V.73A. N4. P.679-689.
Lamprecht L, Zotin A.I. (Eds.) Thermodynamics of Biological Processes. Berlin:
Walter de Gruyter. 1978. 428 p.
Lamprecht L, Zotin A.I. (Eds.) Thermodynamics and Kinetics of Biological
Processes. Walter de Gruyter. 1883. 536 p.
Lamprecht L, Zotin A.I. (Eds.) Thermodynamics and Regulatin of Biological
Processes. Walter de Gruyter. 1985. 573 p.
Lamprecht L, Zotin A.I. (Eds.) Thermodynamics and Pattern Formation in
Biology. Walter de Gruyter.1988. 518 p.
Landauer R. Entropy changes for steady-state fluctuations // J. stat. Phys. 1973.
V.9. N4. P.351-372.
Lasiewski R.C., Dawson W.R. A re-examination of the relation between standard
metabolic rate and body weight in birds // Condor. 1967. V.69. N1. P. 13-
23.
Leach WJ. Oxygen consumption of lampreys, with special reference to
metamorphosis and phylogenetic position // Physiol. Zool. 1946. V.I9.
N3. P.365-374.
Lebsack С.S. Effect of temperature and salinity on the oxygen consumption of the
chiton, Mopalia lignosa II Veliger. 1975. V.18 (Suppl). P.94-97.
Leclerc M., Augarde J. Effects de l'ablation de l'organe axial sur l'activité
respiratoire chez Asterina gibbosa (echinodermes, astérides) // Mar. Biol.
1970. V.6. Nl.P.77-80.
Ledebur J.F. Über die Atmung der Schwämme und Coelenteraten // Ergeb. Biol.
1939. Bd.16. S.262-291.
Le Roy E. Luxidence idaliste et le fait de l'volution. Paris: 1927. 270 p.
Leuschner D. Grundbegriffe der Thermodynamik. Berlin: Akademie Verlag. 1979.
186 S.
Leuschner D. Biomathematics in embryogenesis and the conception of the
phoronomics of the entropy // Biomatrische Z. 1981. Bd.31. H.7. S.l-7.
Leuschner D. Thermodynamik in der Biologie. Berlin: Akademie. 1989. 202 S.
Levine N.D., Corliss J.O., Cox F.E.G., Deroux G., Grain J., Hanigberg B.M.,
Leedale G.F., Loeblich A.R., Lorn J., Lynn D., Merinfeld E.G., Page E.G.,
Poljansky G., Sprague V., Vavra J., Wallace E.G. A newly revised
classificftion of the Protozoa // J. Protozool. 1980. V.27. N.I. P.37-58.
Lewis S.V., Potter I.C. Oxygen consumtion during the metamorphosis of the
parasitic lamprey, Lampetra fluviatilis (L.) and its non-parasitic
derevative, Lampetra planen (Bloch) // J. exp. Biol. 1977. V.69. P.187-
198.
Lighten J.R.B. Ventilation in namib desert tenebrionid beetles: mass scaling and
evidence of a novel quantized flutter-phase // J. exp. Biol. 1991. V.I59.
P.249-268.
Lighten J.R.B., Bartholomew G.A., Feener D.H. Energetics of locomotion and
load carriage and a model of the energy cost of foraging in the leaf-cutting
ant Atta colombica Guer. // Physiol. Zool. 1987. V.60. N5. P.524-537.
Lilly G.R. The influence of diet on the oxygen uptake of the sea urchin,
Triphenstes ventricosus and Strongylocentrotus droebachiensis II Сотр.
Biochem. Physiol. 1979. V.62A. N2. P.463-469.
Lindstedt S.L., Calder W.A. Body size and longevity in birds // Condor. 1976.
V.78. Nl.P.91-94.
Loewy A. Physiologie des Höhenklimas. Berlin: Springer. 1932. 316 S.
L0vtrup S. Victims of ambition: Comments on the Wiley and Brooks approach to
evolution // Syst. Zool. 1983. V.32. N1. P.90-96.
Luebbert S.J., McGregor L.E., Roberts J.C. Temperature acclimation in the
Mongolian gerbil (Meriones unguiculatus): Change in metabolic rate and
the response to norepinephrine // Сотр. Biochem. Physiol. 1979. V.63.
Nl.P.169-175.
Lurie D., Wagensberg J. Entropy balance in biological development and heat
dissipation in embryogenesis // J.Non-Equilib. Thermodyn. 1979a. V.4.
N2. P.127-130.
Lurie D., Wagensberg J. Non-equilibrium thermodynamics and biological growth
and development // J. theoret. Biol. 1979b. V.78. N2. P.241-250.
Lustick S. Bird energetics: Effects of artificial radiation // Science. 1969. V.I63.
P.387-390.
Luxton M. Studies on the orbited mites of a Danish beech wood soil. II. Biomass,
calorimetry and respirometry // Pedobiologia. 1975. V. 15. N 3. P. 161-
200.
Lwoff A. L'e'volution physiologique: Etude des pertes de fonctions chez les
microorganismes. P.: Hermann. 1944. 308 p.
MacKay W.P. An altitudinal comparison of oxygen consumption rates in three
species Pogonomyrmex harvegter ants (Hymtnoptera: Formicidae) //
Physiol. Zool. 1982. V.55. N4. P.367-377.
MacNally R.C. An analysis of factors affecting metabolic rates of two species of
Ranidella (Anura) // Сотр. Biochem. Physiol. 1981. V.69A. N4. P.731-
737.
MacNally R., Young D. Song energetics of the bladder cicada, Cystosoma
saundersii IIJ. exp. Biol. 1981. V.90. P. 185-196.
Maddock L., Young J.Z. Quantitative differences among the brains of cephalopods
// J. Zool. 1987. V.212. N4. P.739-767.
Magniez P., Feral J.-P. The effect of somatic and gonadal size on the rate of
oxygen consumption in the subantarctic echinoid Arbatus cordatus
(Echinodermata) from Kerguelen // Сотр. Biochem. Physiol. 1988.
V.90A. N3. P.429-434.
Maginniss L.A., Wells MJ. The oxygen consumption of Octopus cyanea II J. exp.
Biol. 1969. V.51. N3. P.607-613.
Mallouk R.S. Longevity in vertebrates is proportional to relative brain weight //
Fed. Proc. 1975. V.34, N12. P.2102-2103.
Maloeuf N.S.R. Studies on the respiration (and osmoregulation) of animals. I.
Aquatic animals without oxygen transportes in their internal medium // Z.
vergl. Physiol. 1937. V.25. N1. P. 1-28.
Mangum C.P. Studies on speciation in maldonid polychaetes of the North
American Atlantic coast. III. Intraspecific and interspecific divergence in
oxygen consumption // Сотр. Biochem. Physiol. 1974. V. 10. N.4. P.
335-349.
Marder J. Temperature regulation in the bedoin fowl (Callus domesticus). II
Physiol. Zool. 1973. V. 46. N 3. P. 208-217.
Martin R.D. Relative brain size and basal metabolic rate in terrestrial vertebrates //
Nature. 1981. V.293. P.57-60.
McClendon J.F. The direct and indirect calorimetry of Cassiopea xamachana. The
effect of stretching on the rate of the nerve impulse // J. biol. Chem. 1917.
V.32. P.275-296.
McPherson B.F. Feeding and oxygen uptake of the tropical sea urchin Eucidaris
tribuloides (Lamarck) // Biol. Bull. 1968. V.135. N2. P.308-321.
Meda E. Basal metabolism of man in mountains // Arch. Physiol. 1955. V.551.
N1/2. P.144-151.
Moritz R.F.A., Southwick E.E. Bees as Superorganisms. An Evolutionary Reality.
Berlin: Springer. 1992. 395 p.
Münz F.W., Morris R.W. Metabolic rate of the hadfish, Eptatretus stoutii
(Lockington, 1878) // Сотр. Biochem. Physiol. 1965. V.16. N1. P.l-16.
Murdoch R.C., Shumway S.E. Oxygen consumption in six species of chitons in
relation to their position on shore // Ophelia. 1980. V.19. N2. P.127-144.
Mûrie M. Metabolic characteristics of mountain, desert and coastal population of
Peromyscus II Ecology. 1961. V.42. N.4. P.723-740.
Nageli C. Mechanisch-physiologische Theorie der Abstammungslehre. München.
1884.
Neal C.M., Lustick S.I. Energy budget of the eastern chipmunk (Tamias striatus}:
Artificial radiation // Сотр. Biochem. Physiol. 1975. V.50A. N2. P.233-
236.
Nicolis G. Stability and dissipative structures in open systems far from equilibrium
// Adv. chem. Phys. 1971. V.19. P.209-324.
Nicolis G., Babloyantz A. Fluctuations in open systems // J. chem. Phys. 1969.
V.51.N6. P.2632-2637.
Nicolis G., Prigogine I. Fluctuations in nonequilibrium systems // Proc. nat. Acad.
Sei. USA. 1971. V.68. N9. P.2102-2107.
Nielsen M.G., Jensen T.F., Holm-Jensen I. Effect of load carriage on the
respiratory metabolism of running worker ants of Camponotus
herculeanus (Formicidae) // Oikos. 1982. V.39. N2. P.137-142.
Nival P., Nival S., Palazzoli I. Données sur la respiration de différents organismes
communs dans le plancton de Villefranche-sur-Ver // Mar. Biol.. 1972.
V.17. Nl.P.63-76.
Noll U.G. Body temperature, oxygen consumption, noradrenaline response and
cardiovascular adaptations in the flying fox, Rousettus aegyptiacus II
Сотр. Biochem. Physiol. 1979. V.63A, N1. P.79-88.
О Dor R.K. Respiratory metabolism and swimming performance of the squid,
Loligo opalescens II Canad. J. Fish. Aquat. Sei. 1982. V.39. N4. P.580-
587.
Onsager L. Reciprocal relations in irreversible processes. I // Phys.Rev. 193la.
V.37. N4. P.405-426.
Onsager L. Reciprocal relations in irreversible processes. II. // Phys.Rev. 1931b.
V.38. N12. P.2265-2279.
Ordy J.M. Neurobiology and aging in nonhuman primates // Neurobiology of
aging / Eds. J.M.Ordy et al. New York: Plenum Press. 1976. P.575-594.
Pasquis P., Lacaisse A., Dejours P. Maximal oxygen uptake if four species of
small mammals // Respirât. Physiol. 1970. V.9. N2. P.298-309.
Patterson J.W., Davies P.M.C. A possible effect of climate on resting metabolic
rate in lizards // Copeia. 1989. N3. P.719-723.
Paul R., Fincke T., Linzen B. Book lung function in arachnids. I. Oxygen uptake
and respiratory quotient during rest, activity and recovery-relations to gas
transport in the haemolymph // J. Сотр. Physiol. 1989. V. 159B. N 4.
P.409-418.
Pearson O.P. The metabolism of humminghbirds // Sei. Amer. 1953. V. 188,
Nl.P.69-72.
Pennycuik P.R. A comparison of the effect of a variety of factors on the metabolic
rate of the mouse // Austral. J. Exp. Biol. Med. Sei. 1967. V.45. N4.
P.331-346.
Pentreath RJ. Respiratory surfaces and respiration in three New Zealand intertidal
ophiuroids // J. Zool. 1971. V.163.N3. P.397-412.
Percy J.A. Thermal adaptation in the borea-arctic echinoid Strongylocentrotus
droebachiensis (O.F.Müller). I. Seasonal aclimatization of respiration //
Physiol. Zool. 1972. V.45. N4. P.277-289.
Phillipson J. Respiratory and the study of energy turnover in natural systems with
particular reference to harvest-spiders (Phalangiidae) // Oikos. 1962. V.13.
N2. P.311-322.
Phillipson J. The use of respiratory data in estimating annual respiratory
metabolism with particular reference to Leiobunum rotundum (Latr.)
(Phalangiidae) // Oikos. 1963. V.14. N2. P.212-223.
Piiper J., Baumgarten D., Meyer M. Effects of hypoxia upon respiration and
circulation in the dogfish Scyliorhinus stellaris II Сотр. Biochem.
Physiol. 1970. V.36. N3. P.513-520.
Popper K.R. Time's arrow and entropy // Nature. 1965. V.207. P.233-234.
Popper K.R. Time's arrow and feeding on negentropy // Nature. 1967a. V.213.
P.320.
Popper K.R. Structural information and the arrow of time // Nature. 1967b. V.214.
P.322.
Porter S. Revised respiration rates for the southern harvester ant, Pogonomyrmex
badius II Сотр. Biochem. Physiol. 1986. V.83A. N.I. P.197-198.
Potter I.C., Rogers M.J. Oxygen consumption in burrowed and unburrowed
ammocoetes of Lampetra planen (Bloch) // Сотр.Biochem. Physiol.
1972. V.41A. N2. P.427-432.
Prestwich K.N. The roles of aerobic anaerobic metabolism in active spider //
Physiol. Zool. 1983. V.56. N1. P.122-132.
Prigogine I. Etude thermodynamique des phnomnes irrversible. P.: Desoer. 1947.
143 p.
Prigogine I. La thermodynamique de la vie // La Recherche. 1972. T.3, N24.
P.547-562.
Prigogine L, Nicolis G., Babloyantz A. Thermodynamics of evolution // Physics
Today. 1972. V.25, N11. P.23-28; V.25, P. 12. P.38-44.
Prinzinger R. Lebensalter und gesamtenergieproduktion beim Vgel // J. Ornithol.
1979. V.120, Nl.P.103-105.
Prinzinger R. The energy cost of life stages in birds // Energy Transformations in
Cells and Organisms / Eds. W.Wieser, E.Gnaiger. New York: Thieme,
1989. P.123-129.
Rajagopal P.K. Respiration of some marine planktonic organisms // Proc. Indian
Acad. Sei. 1962. V.55B, N2. P.76-81.
Redmond J.R., Bourne G.B., Johansen K. Oxygen uptake by Nautilus pampilus II
J. exp. Zool. 1978. V.205. N1. P.45-49.
Reeve M.R., Raymond J.E., Raymond J.K.B. Seasonal biochemical composition
and energy sources of Sagitta hispida // Mar. Biol. 1970. V.6. N4. P.357-
364.
Reichle D.E. Relation of body size to food intake oxygen consumption, and trace
element metabolism in forest floor arthropods // Ecology. 1968. V. 49. N
3.P. 538.
Reiss J.O. The meaning of developmental time: a metric for comparative
embryology // Amer. Natur. 1989. V.2. P.170-189.
Rench B. Neuere Probleme der Abstammungslehre. Die Transspezifische
Evolution. Stuttgart. 1954. 468 S.
Rench B. Evolution Above the Species Level. New York: Columbia Univ. Press.
1960.419p.
Richet Ch. Recherches de calorimetrie // Arch. Physiol. norm. Pathol. 1885. N.7.
P.237-291;N.8. P.450-497.
Richet Ch. De la mesure des combustions respiratoires chez les mammifères //
Arch. Physiol. norm. Pathol. 1891. V.3. N.I. P.74-76.
Rieck A.F., Belli J.A., Blaskovics M.E. Oxygen consumption of whole animal and
tissues in temperature acclimated amphibians // Proc. Soc. Exp. Biol. Med.
1960. V.103.N2. P.436-439.
Robbins B.A. Aerial and aquatic respiration in the chitons Nuttallina californica
and Tonicella lineata. Veliger. 1975. V.18 (Suppl.). P.98-102.
Roberts J.C., Chaffee R.R.J. Metabolic and biochemical aspects of heat
acclimation in the deer mouse, Peromyscus maniculatus sonoriensis //
Сотр. Biochem. Physiol. 1976. V.53A. N4. P.367-373.
Roberts J.C., Hock R.J., Smith R.E. Seasonal metabolic responses of deer mice
(Peromyscus) to temperature and altitude // Fed. Proc. 1966. V.25. N4.
P.1275-1283.
Robertson D.A. Volume changes and oxygen extraction efficiences in the
holothurian, Stichopus mollis (Hutton) // Сотр. Biochem. Physiol. 1972.
V.43A. N4. P.795-800.
Rubner M. Über den Einfluss der Körpergrösse auf Stoff- und Kraftwechel //
Ztschr. Biol. 1883. Bd. 19. S. 535-562.
Rubner M. Die Gesetze der Energieverbrauchs bei der Ernährung.
Leipzig.Deiticke. 1902. 426 S.
Rubner M. Das Problem der Lebensdauer und seine Beziehungen zu Wachstum
und Ernhrung. Mnchen: Oldenburg, 1908a. 208 S.
Rubner M. Problem des Wachstums und den Lebensdaner // Gesellschaft für inner
Medizin und Kinderheikunde. Wien. 1908b. Bd.7. S.58-72.
Sacher G. A. Relation of lifespan to brain weight and body weight in mammals //
Ciba Foundation Colloquia on Ageing. V.5. The Lifespan of Animals /
Eds. G.E.Wolstenholme, M.O'Connor. London: Churchill, 1959. P. 115-
141.
Sacher G.A. Maturation and longevity in relation to cranial capacity in hominid
evolution // Primate Functional Morphology and Evolution / Eds.
R.H.Tuttle. Hague-Paris: Mounton. 1975. P.417-441.
Sacher G.A. Evaluation of the entropy and information terms governing
mammalian longevity // Cellular Ageing: Concepts and Mechanisms. Part
I. General Concepts. Mechanisms I: Fidelity of Information Flow / Ed.
R.G.Culter. Basel: Karger. 1976. P.69-82.
Sacher G.A. Longevity, aging, and death: an evolutionary perspective //
Gerontologist. 1978. V.18. N1. P.I 12-119.
Sagawa S., Hanion R.T. Oxygen consumption and ammonia rates in Octopus
maya, Loligo forbesi and Lolliguncula brevis (Mollusca: Cephalopoda) //
Mar. Behav. Physiol. 1988. V.13. N4. P.389-400.
Saroja K. Studies on the oxygen consumption in tropical poikilotherms. 2. Oxygen
consumption in relation to body size and temperature in the earthworm
Megascolex mauritii when kept submerged in water // Proc. Indian Acad.
Sei. 1959. V. 49B. N.3. P. 183-193.
Saroja K. Seasonal acclimatization of oxygen consumption to temperature in a
tropical poikilotherm, the earthwarm, Megascolex mauritii II Nature. 1961.
V.190. P.930-931.
Saroja K. Oxygen consumption of the worm Octochaetona s errata as a function of
size and temperature in aquatic and aerial media // Сотр. Biochem.
Physiol. 1964. V. 12. N.I. P. 47-53.
Sassaman C., Mangum C.P. Gas exchange in a cerianthid // J. exp. Zool. 1974.
V.188. N3. P.297-306.
Saunders P.T., Ho M.W. On the increase in complexity in evolution. The relativity
off complexity and the principle of the minimum increase // J. theoret.
Biol. 1981. V.90. N.4. P.515-530.
Savage D.E., Rüssel D.E. Mammalian Paleofauna of the World. Mass.: Adison,
Wesley. 1983. 432 p.
Scheck S.H. A comparison of thermorégulation and evaporation water loss in two
hispid cotton rat, Sigmodon hispidus texianus, from northern Kansas and
south-central Texas // Ecology. 1982. V.63. N2. P.361-369.
Schölander P.E., Flagg W., Walters V., Irving L. Climatic adaptation in arctic and
tropical poikilotherms // Physiol. Zool. 1953. V.26. N1. P.67-92.
Schölander P.E., Hock R., Walters V., Johnson F., Irving L. Heat regulation in
some arctic and tropical mammals and birds // Biol. Bull. 1950. V.99. N2.
P.237-258.
Schwemmler W. Symbiogenesis. A Macro-Mechanism of Evolution. Btrlin: de
Gruyter, 1989. 225 p.
Sewertzoff A.N. Morphologische Gesetzmssigkeiten der Evolutien. Jena: Fischer.
1931. 371 S.
Shick J.M., Brown W.I., Dolliver E.G., Kayar S.R. Oxygen uptake in sea
anemones: effects of expansion, contraction, and exposure to air and the
limitations of diffusion // Physiol. Zool. 1979. V.52. N1. P.50-62.
Shield J.W., Bently P.J. Respiration of some urodele and anuran Amphibia. I. In
water, role of the skin and lungs // Сотр. Biochem. Physiol. 1973. V.46A,
Nl,P.17-28..
Sidell B.D. Turnover of cytochrome с in skeletal muscle of green sunfish
(Lepomas cyanollus R.) during thermal acclimation // J. exp. Zool. 1977.
V.199. N2. P.233-250.
Silverthorn S.U. Respiration in eyestalkless Uca (Crustacea: Decapoda) acclimated
to two temperatures // Сотр. Biochem. Physiol. 1973. V.45A. N2. P.417-
420.
Simpson G.G. The Major Features of Evolution. New York: Columbia Univ.
Press. 1953. 434 p.
Simpson G.G. The Meaning of Evolution. New Haven (Conn.): Yale Univ. Press.
Chapter 15. 1967. 384 p.
Sisak M.M., Sander F. Respiratory behaviour of the Western Atlantic
holothuroidian (Echinodermata) Holothuria glaberrima (Selenka) at
various salinities, temperatures and oxygen tensions // Сотр. Biochem.
Physiol. 1985. V.80A. N1. P.25-29.
Smith G.C. Ecological energetics of three species of ectothermic vertebrates //
Ecology. 1976. V.57. N2. P.252-264.
Smith K.L. Metabolism of two dominant epibenthic echinoderms measured at
Bathyal Depns in the Santa Catalina Basin // Mar. Biol. 1983. V.72. N3.
P.249-256.
Smith K.L., Hessler R.R. Respiration of benthopelagic fishes: in situ
measurements at 1230 meters // Science. 1974. V.I84. P.72-73.
Smyth J.D. The Physiology of Cestodes. Edinburgh: Oliver, Boyd.1966. 279 p.
Southwick E.E. Allometric relations, metabolism and heat conductance in clusters
of honey bees at cool temperatures // J. сотр. Physiol. 1985. V.156B. N1.
P.143-149.
Southwick E.E. Thermorégulation in honey-bee colonies // Africanized honey bee
and bee mites / Eds. G.Needham et al. New York: Wiley. 1988. P.223-
236.
Southwick E.E. How will Africanized honey bees behave? // Amer. Bee J. 1990.
V.130. N3.P.200-201.
Steen J.B. Comparative aspects of the respiratory gas exchange of sea urchin //
Acta Physiol. Scand. 1965. V.63. N.l/2. P.164-170.
Steigen A.L. Energy metabolism in the wolf spider Pardosa palustris (L.)
(Araneae, Lycosidae) from Hardangervidda, Southern Norway //
Norwegian J. Entomol. 1976. V. 23. N 1. P. 51-60.
Stephan H., Andy OJ. Quantitative comparisons of brain structures from
insectivores to primates // Amer. Zool. 1964. V.4. N1. P.59-74.
Stephan H., Bauchot R., Andy OJ. Data on size of the brain and of various part in
insectivores and primates // The Primate Brain / Eds. C.R.Noback,
W.Montagna. New York: Appeton. 1970. P.289-297.
Stickney J.C., Van Liere EJ. Acclimatization to low oxygen tension // Physiol.
Rev. 1953. V.33. N1. P.13-34.
Studier E.H., O'Farrell M.J. Biology of Myotis thysanodes and M. lacifugus
(Chiroptera: Vespertilionidae). III. Metabolism, heart rate, evaporative
water loss and general energetics // Сотр. Biochem. Physiol. 1976.
V.54A. N4. PA23-432.
Süess E. Die Entstehung der Alpen. Wien: 1875. 168 S.
Swensen R. Emergent attractors to a theory of general evolution // Syst. Res. 1989.
V.6.N3. P. 187-197.
Swensen R. Order, evolution, and natural law: fundamental relations in complex
system theory // Cybernetics and Applied Systems / Ed. C.V.Negoita.
N.Y.: Dekker. 1992. P. 125-148.
Swensen R. Thermodynamics, evolution, and behavior // The Encyclopedia of
Comparative Psychology / Eds. G.Greenberg, M.Haraway. N.Y.: Garland.
1997. P. 217-228.
Taigen T.L. Activity metabolism of anuran amphibians: implications for the origin
of endothermy // Amer. Nat. 1983. V.121. N1. P.94-109.
Templeton J.R. Reptiles // Comparative Physiology of Thermorégulation / Ed.
G.C.Whittow. N.Y.: Acad. Press. 1970.. New York: Acad.Press, 1970.
V.l.P.167-221.
Tilbrook P.J., Block W. Oxygen uptake in an Antarctic collembole Cryptopysus
antarcticus II Oikos. 1972. V.23. N3. P.313-317.
Tolmasoff J.M., Ono T., Culter R.G. Superoxide dismutase: correlation with life-span and specific metabolic rate in primate species // Proc. nat.
Acad.Sci.USA. 1980. V.77. N5. P.2777-2781.
Torrance J.D., Lenfant C., Cruz J., Marticorena E. Oxygen transport mtchanisms
in residents at high altitude// Respirât. Physiol. 1970/1971. V.U. N1. P.l-
15.
Touloukian Y.S. The Concept of Entropy in Communication, Living Organisms
and Thermodynamics. Lafayette (Ind.): Purdae Univ.Press. 1956. 66 p.
Trueman E.R., Bone Q., Braconnot J.C. Oxygen consumption in swimming salps
(Tunicata: Thaliacea) // J. exp. Biol. 1984. V.I 10. P.323-327.
Tykodi RJ. Thermodynamics of steady states: a weak entropy-production
principle // Physica. 1974. V.72. N2. P.341-354.
Ulanowicz R.E. Growth and Development Ecosystems Phenomenology. New
York: Springer. 1986. 203 p.
Ulbricht R. J. Effect of temperature acclimation on the metabolic rate of sea urchin
// Mar. Biol., Berlin. 1973a. V.19. N4. P.273-277.
Ulbricht RJ. The influence of temperature acclimation upon the metabolic rate of
the purple sea urchin, Strongylocentrotus purpuratus: alternate
interpretations // Сотр. Biochem. Physiol. 1973b. V. 45A. N2. P.677-681.
Ulbricht R.J., Pritchard A.W. Effect of temperature on the metabolic rate of sea
urchins // Biol. Bull. 1972. V.142. N1. P.178-185.
Ullrick W.C., Whitehorn W.V., Brennan B.B., Krone J.G. Tissue respiration of
rate acclimation tolow barometric pressure // J. appl. Physiol. 1956. V.9.
N.l.P.49-52.
Ultsch G.R. Gas exchange and metabolism in the Sirenidae (Amphibia: Caudata).
I. Oxygen consumption of submerged sirenids as a function of body size
and respiratory surface area // Сотр. Biochem. Physiol. 1974. 47A. N2.
485-498.
Van Dam L. On the respiration in scallops (Lamellibranchiata) // Biol. Bull. 1954.
V.107. N2. P. 192-202.
Vandel A. L'Homme et 1'evolution. P.: Gallimard. 1949. 201 p.
Vandel A. La genèse du vivant // Ann. biol. 1967. V.6. N9/10. P.579-588.
Van Hook R.I. Energy and nutrient dynamics of spiders and orthopteran
population in a grass land ecosystems // Ecol. Monogr. 1971. V. 41. N 1.
P. 1-26.
Vernberg FJ. Studies on the physiological variation between tropical and
temperate zone fidder crabs of the genus Uta. 2. Oxygen consumption of
whole organism//Biol. Bull. 1959. V.I 17. N1. P.163-184.
Vernberg W.B. Platyhelminthes: respiratory metabolism // Chemical Zoology. /
Eds. M.Florkin, B.T.Scheer. New York: Acad.Press. 1968. V.2. P.359-
394.
Vernberg W.B., Hunter W.S. Studies on oxygen consumption in digjnetic
trematodes. V. The influence of temperature on three species of adult
trematodes // Exp. Parasit. 1961. V.U. Nl.P.34-38.
Vinegar A., Hutchison V.H., Dowling H.G. Metabolism, energetics, and
thermorégulation during brooding of snakes of the genus Python (Reptilia,
Boidae) //Zoologica. 1970. V.55. N2. P.19-48.
Visinescu N. The particularités of thermorégulation and their seasonal variations in
Clethrionomys glareolus Schreb. (1780) and Apodemus sylvaticus L.
(1758) // Rev. Roum. Biol. Zool. 1967. V.12. N2. P.127-137.
Viswanadham C.R. Entropy, evolution and living systems // Nature. 1968. V.219.
P.653.
Vladimirova I.G., Alekseeva T.A., Lebedeva A.A. Oxygen consumption of some
Amphibian sprcies // Advances in Amphibian Biology in the Former
Soviet Union / Eds. S.L.Kuzmin. Sofia-Moscow: Pensoft. 1997. P. 157-
162.
Vojta G. Hamiltonian formalism in thermodynamic theory of irreversible
processes in continuous systems // Acta chim. Acad. Sei. Hungr. 1967.
V.54. Nl.P.55-64.
Waddington C.H. The Strategy of the genes. L.: Allen, Unwin.1957. 262 p.
Walkey M., Davies P.S. Effect of pH on oxygen consumption of the cestode
Schistocephalus solidus II Exp.Parasit. 1968. V.22. N2. P.201-206.
Wallace J.C. Observation on the relationship between the food onsumption and
metabolic rate of Blennius pholis L. // Сотр. biochem. Physiol. 1973.
V.45A. N2. P.293-306.
Wangsness R.K. Minimum entropy production and logarithmic rate equations //
Physica. 1975. V.79A. N5. P.543-557.
Warnock R.E., Liddell W.D. Oxygen consumption in two shallow-water comatulid
crinoids // J. exp. mar. Biol. Ecol. 1985. V.91. N1/2. P.169-182.
Weathers W.W. Climatic adaptation in avian standard metabolic rate // Oecologia.
1979. V.42. Nl.P.81-89.
Webb N.R. Oxygen consumption and population metabolism of some
mesostigmated mites (Acari: Mesostigmata) // Pedobiologia. 1970. V.10.
N6. P.447-456.
Webber D.M., О Dor R.K. Monitoring the metabolic rate and activity of free-swimming squid with telemetred jet pressure // J. exp. Biol. 1986. V.I26.
P.205-224.
Webster S.K. Oxygen consumption in exhinoderms from several geographical
locations, with particular reference to the Echinoidea // Biol. Bull. 1975.
V.148. Nl.P.157-164.
Webster S.K., Giese A.C. Oxygen consumption of the purple sea urchin with
special reference to the reproductive cycle // Biol. Bull. 1975. V.148. N1.
P.165-180.
Wells M.J. The brain and behavior of cephalopods // The Physiology of Mollusca /
Eds. K.M.Wilbur, C.M.Younge. New York: Acad. Press. 1966. V.2..
Wells M.J., Hanion R.T., Lee P.O., Dimarco P.P. Respiratory and cardiac
performance in Lolliguncula brevic (Ctphalopoda, Myopsida): The effect
of activity, temperature and hypoxia // J. exp. Biol. 1988. V.138. P. 17-36.
Wells M.J., О Dor R.K., Mangold K., Wells J. Feeding and metabolic rate in
Octopus JI Mar. Behav. Physiol. 1983. V.9. N4. P.289-303.
Wells M.J., Wells J. The circulatory response to acute hypoxia in Octopus II J.
exp. Biol. 1983. V.104. P.59-71.
Wells M.J., Wells J. Ventilation and oxygen uptake my Nautilus II J. exp. Biol.
1985. V.118. P.297-312.
Wells N.A. Variations in the respiratory metabolism of the pacific killfish
Fundulus parvipinnis due to size, season and continued constant
temperature // Physiol. Zool. 1935. V.8. N2. P.318-336.
Whitford W.G., Hutchison V.H. Body size and metabolic rate in salamanders //
Physiol.Zool. 1967. V.40. N2. P.127-133.
Whitney R.J. The relation of animal size to oxygen consumption in some
freshwater Turbellarian worms // J. exp. Biol. 1942. V.19. N2. P. 168-175.
Wicken J.S. The generation of complexity in evolution: A thermodynamic and
information-theoretical discussion // J. theoret. Biol. 1979. V.77. N3.
P.349-365.
Wicken J.S. A thermodynamic theory of evolution // J. theoret. Biol. 1980. V.87.
Nl.P.9-23.
Wicken J.S. Thermodynamics and the conceptual structure of evolutionary theory
// J. theoret. Biol. 1985. V.I 17. N3. P.263-283.
Wicken J.S. Entropy and evolution: Ground rules for discourse // Syst. Zool. 1986.
V.35. Nl.P.22-36.
Wicken J.S. Evolution, Thermodynamics, and Information. Oxford: Univ. Press.
1987. 243 p.
Wiegert R.G. Population energetics of meadow spittlebugs (Philaenus spumarius
L.) as affected by migration and habitat // Ecol. Monogr. 1964. V.34. N3.
P.217-241.
Wieser W. A distinction must be made between the ontogeny and the phylogeny of
metabolism in order to understand the mass exponent of energy
metabolism//Respir. Physiol. 1984. V.55. N1. P.l-9.
Wigand A. Der Darwinismus und die Naturforschung Newtons and Cuvier.
Braunschweig, 1874.
Wigglesworth V.B. The Principles of Insect Physiology. London: Methuen. 1953.
546 p.
Wiley E.O., Brooks D.R. Vistims of history - A nonequilibrium approach to
evolution // Syst. Zool. 1982. V.31. N . P. 1-24.
Wiley E.O., Brooks D.R. Nonequilibrium thermodynamics and evolution: A
response to Lovtrup // Syst. Zool. 1983. V.32. P.209-219.
Wilkie D.R. Thermodynamics and the interpretation of biological heat
measurements // Progr. Biophys. Biophys. Chem. 1960. P.259-298.
Willems N.J., Armitage K.B. Thermorégulation and water requirements in
semiarid and montane populations of the least chipmunk, Eutamius
minimus. I. Metabolic rate and body temperature // Сотр. Biochem.
Physiol. 1975. V.51A. N.4. P.717-722.
Wilson J.A. // Increasing entropy of biological systems // Nature. 1968a. V.219.
P.534-535.
Wilson J.A. Entropy, not negentropy // Nature. 1968b. V.219. P.535-536.
Wilson K.J. The relationships of maximum and resting oxygen consumption and
heart rates to weight in reptiles of the order Squamata // Copeia. 1974.
V.3. P.781-785.
Wilson K.J., Lee A.K. Changes in oxygen consumption and heart-rate activity in
body temperature in the tuatara, Sphendon punctatum II Сотр. Biochem.
Physiol. 1970. V.33. N2. P.311-322.
Withers P.C., Hillman S.S. Oxygen consumption of Amphiuma means during
forced activity and recovery // Сотр. Biochem. Physiol., 1981, V.69A.
N1, P.141-144.
Wood Т.О., Lawton J.H. Experimental studies on the respiratory rates of mites
(Acari) from beech-woodland leaf litter // Oecologia. 1973. V.I2. N2.
P.169-191.
Woolhause H.W. Negentropy, information and the feeding of organisms // Nature.
1967a. V.213. P.952.
Woolhause H.W. Entropy and evolution // Nature. 1967b. V.216. P.200.
Wyles J.S., Kunkel J.G., Wilson A.C. Birds, behavior, and anatomical evolution //
Proc. Nat. Acad. Sei. USA. 1983. V.80. N14. P.4394-4397.
Yamaguchi M. Effect of elevated temperature on the metabolic activity of the
coral reef asteroid Acanthaster planci (L.) // Pacific Sei. 1974. V.28. N2.
P.139-146.
Yarbrough Ch.G. The influenceb of distribution and ecology on the
thermorégulation of small birds // Сотр. Biochem. Physiol. 1971. V.39A.
N2. P.235-266.
Yousef M.K., Dill D.B. Responses of Merriam's kangaroo rat to heat // Physiol.
Zool. 1971. V.44. N1. P.33-39.
Zar J.H. Standard metabolism comparisons between orders of birds // Condor.
1968. V.70. N3. P.278-279.
Zebe E.G. Über den Stoffwechsel der Lepidopteren // Z. vergl. Physiol. 1954.
Bd.36. H.3. S.290-317
Zepelin H., Rechtschaffen A. Mammalian sleep, longevity, and energy metabolism
// Brain Behav. EV. 1974. V.10. N6. P.425-470.
Zeuthen E. Oxygen uptake as related to body size in jrganisms // Quart.Rev.Biol.
1953. V.28. Nl.P.1-12.
Zotin A.A., Lamprecht I, Zotin A.I. Bioenergetic progress and heat barriers. // J.
Soc. Evol. Systems. 2000. (in press).
Zotin A.A., Zotin A.I. Thermodynamic basis of developmental processes // J.Non-
Equilibr.Thermodyn. 1996. V.21. N4. P.307-320.
Zotin A.I. Thermodynamic Bases of Developmental Processes: Physiological
Reactions and Adaptations. В.: Walter de Gruyter. 1990. 293 p.
Zotin A.I. Stability and reliability of ontogenesis // Thermochim. Acta. 1995.
V.251.P.363-369.
Zotin A.I., Lamprecht I. Aspects of bioenergetics and civilizaton // J. theoret. Biol.
1996. V.180. N2. P.207-214.
Zotin A.I., Zotina R.S. Thermodynamic aspects of developmental biology. // J.
Theoret. Biol. 1967. V. 17. N 1. P. 57-75.
Zotin A.I., Zotina R.S. Principle of least dissipation of energy as a basis of
thermodynamic theory of development and growth // First Europ. Biophys.
Congress / Eds. T.Broda, A.Locker. Vien: Springer. 1971. V.6. P.9-12.
РИСУНКИ (1-29):
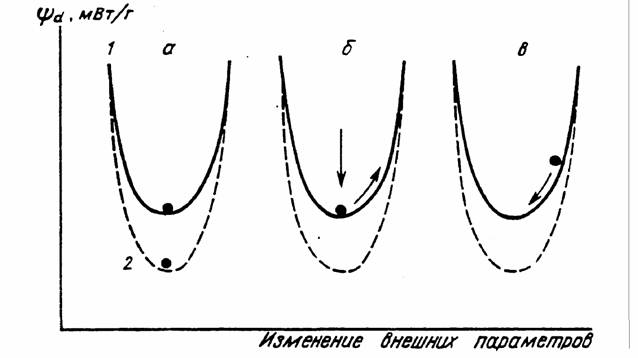
|
| Рис.1. Схема, иллюстрирующая принцип наименьшей диссипации энергии для обычной (1) и организованной (2) систем (Зотин,1988). |
| Шарик на дне чаши (а) соответствует стационарному состоянию системы с минимальной величиной функции внешней диссипации. Стрелками показаны внешнее воздействие на систему, ее уклонение от стационарного состояния (б) и релаксация системы в начальное состояние (в) после прекращения внешнего воздействия. |
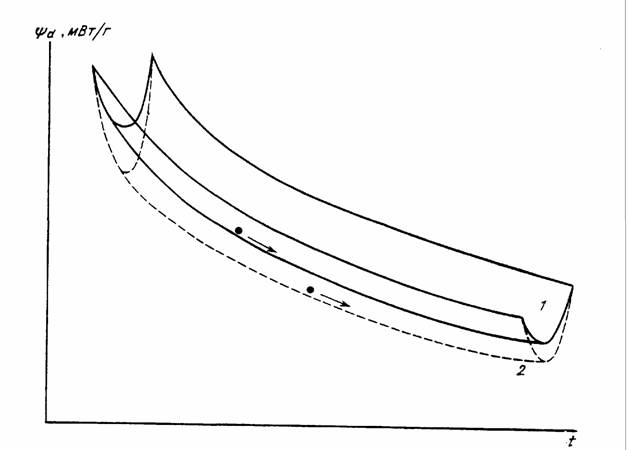
|
| Рис.2. Схема, иллюстрирующая интегральный принцип наименьшей диссипации энергии для обычной (1) и организованной (2) систем (Зотин, Зотина,1993). |
| Креод соответствует наиболее устойчивому стационарному процессу - шарик катится по дну желоба. |
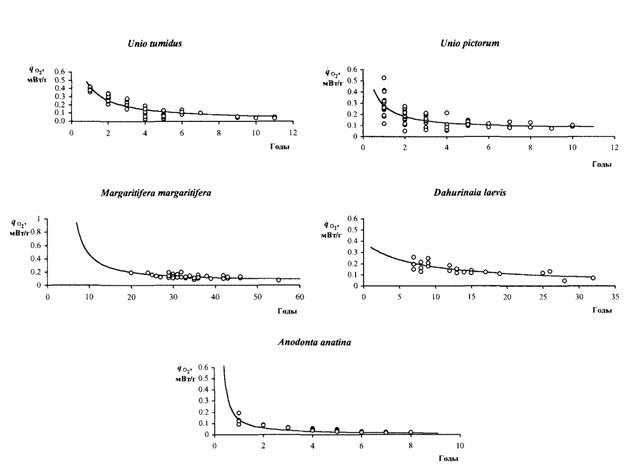
|
| Рис.3. Кинетика изменения интенсивности потребления кислорода различными видами пресноводных двустворчатых моллюсков. |
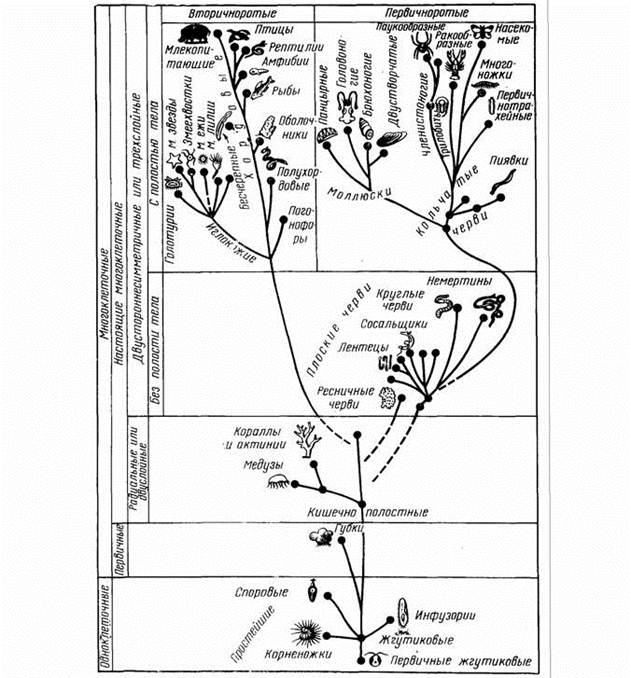
|
| Рис.4. Древо животного мира (Тимофеев-Ресовский и др., 1969). |
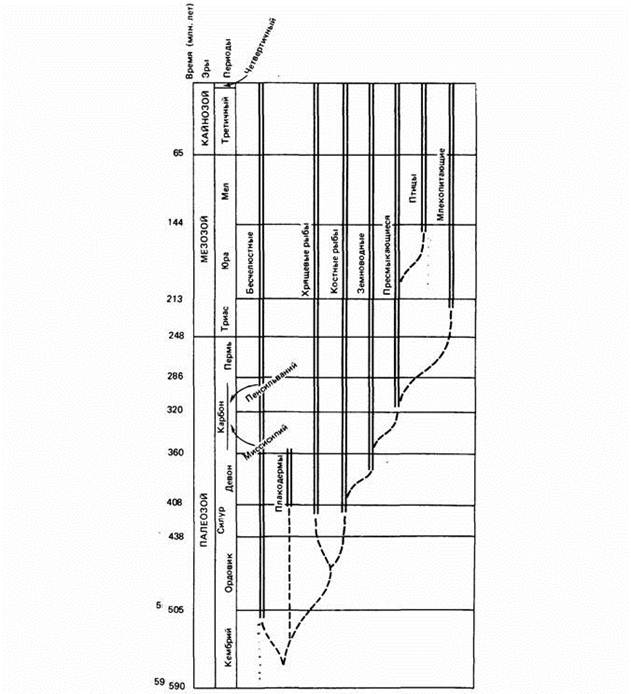
|
| Рис.5. Время существования разных классов позвоночных животных и их предполагаемое родство (Кэрролл, 1992). |
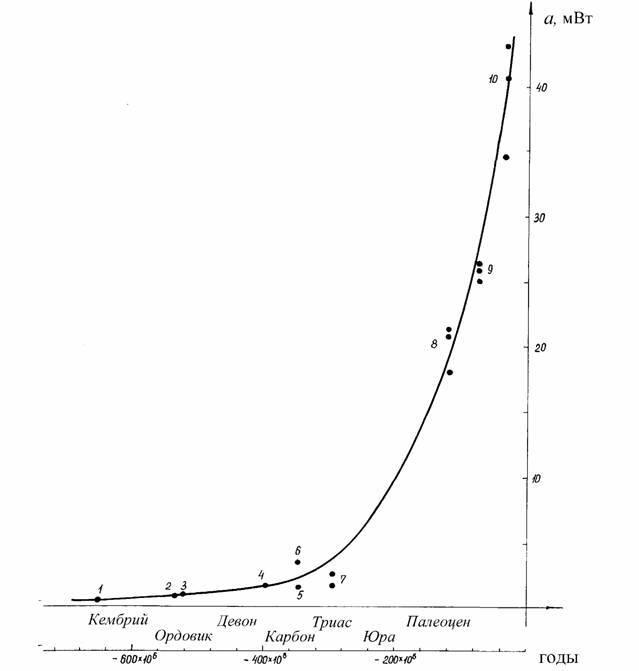
|
| Рис.6. Изменение сопоставимого стандартного обмена (коэффициента а) в процессе биологической эволюции в разных группах животных (Zotin, 1990): 1 - Coelenterata; 2 - Crustacea; 3 - Mollusca; 4 - Teleostei; 5 - Amphibia; 6 - Insecta; 7 - Reptilia; 8 - Mammalia; 9 - Aves; 10 - Passeriformes. |
|
| Рис.7. Видоизмененная схема биологического прогресса (Северцов,1934). Обозначения: ар - ароморфозы; ал - алломорфозы; ка - катаморфозы. |
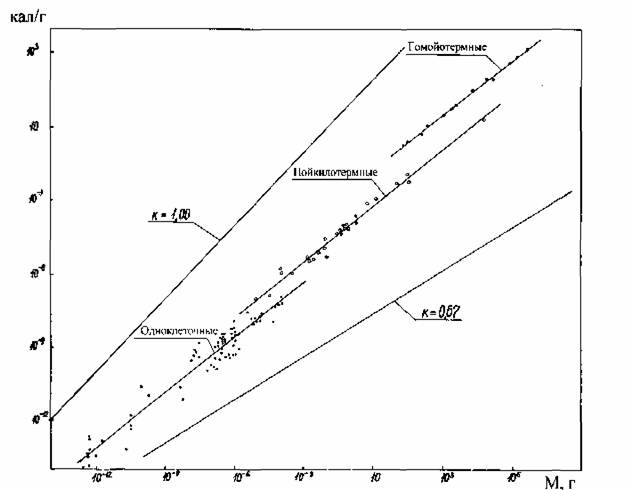
|
| Рис.8. Зависимость стандартного обмена от массы тела у одноклеточных, пойкилотермных и гомойотермных животных (Hemmingsen,1970). |
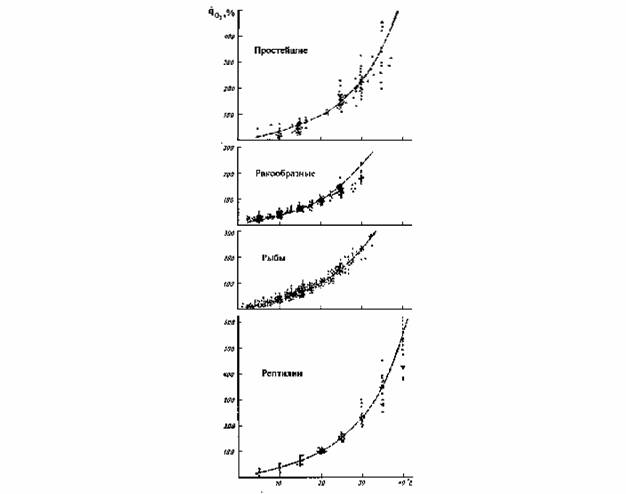
|
| Рис.9. Зависимость потребления кислорода от температуры среды у пойкилотермных животных (Зотин, 1988). Дыхание выражено в % от потребления кислорода при 20°С, принятого за 100%. |
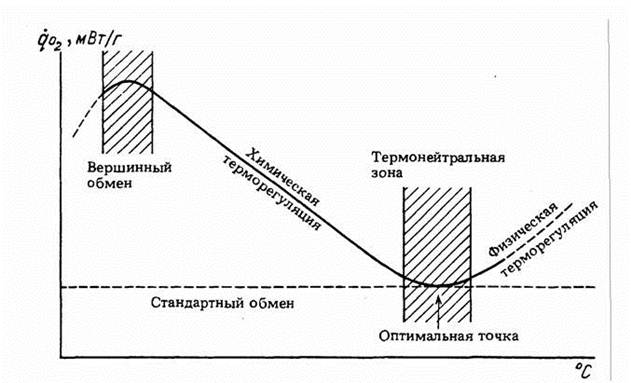
|
| Рис. 10. Схема кривой терморегуляции у гомойотермных животных (Зотин, 1988) |
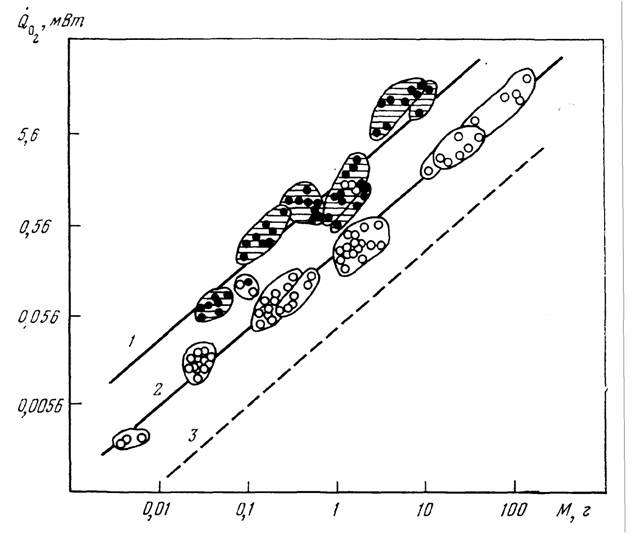
|
| Рис.11. Скорость потребления кислорода ракообразными: тропические виды (1) при 30°С; арктические виды (2) при 0°С (Schölander et al., 1953). Пересчет данных о дыхании тропических видов с помощью кривой Крога на температуру 0°С показан пунктирной прямой (3). |
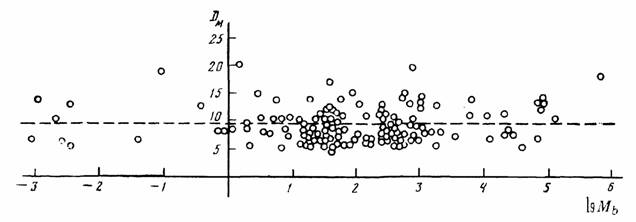
|
Рис.12. Зависимость метаболитического диапазона (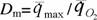 ) от массы тела (Мb, г) животных (Донцова, Зотин, 1980). ) от массы тела (Мb, г) животных (Донцова, Зотин, 1980).
|
|
| Рис.13 Зависимость скорости потребления кислорода при 20°С от массы тела у простейших (Владимирова, Зотин,1983). Обозначения: А - подтип Mastigophora; Б - подтип Sarcodina; В - тип Ciliophora; Г - тип Apicomplexa; 1 - эндогенное дыхание свободноживущих форм; 2 - дыхание на субстратах свободноживущих форм; 3 - дыхание на субстратах паразитических форм. |
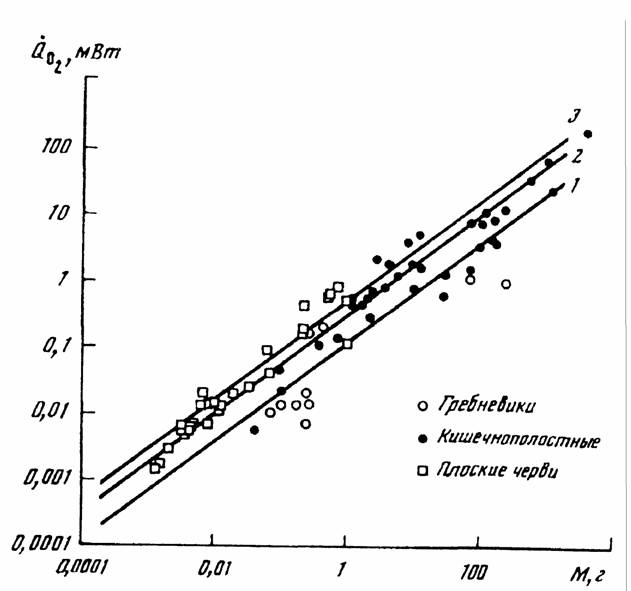
|
| Рис.14. Зависимость скорости потребления кислорода при 20°С от массы тела у гребневиков (1), кишечнополостных (2) и плоских червей (3). |
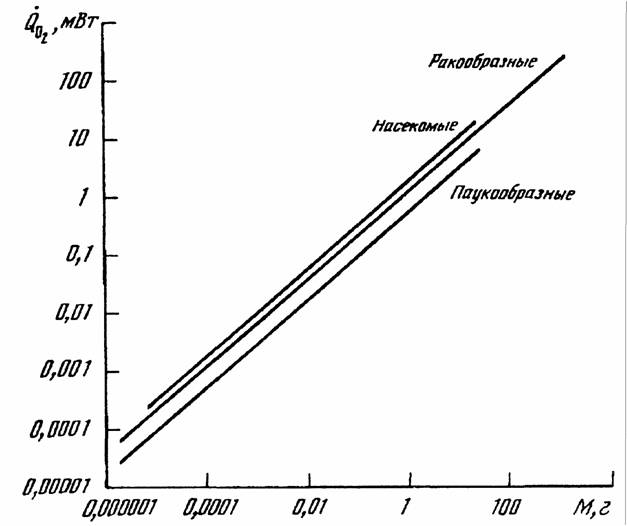
|
| Рис.15. Зависимость скорости потребления кислорода при 20°С от массы тела в типе членистоногих. |
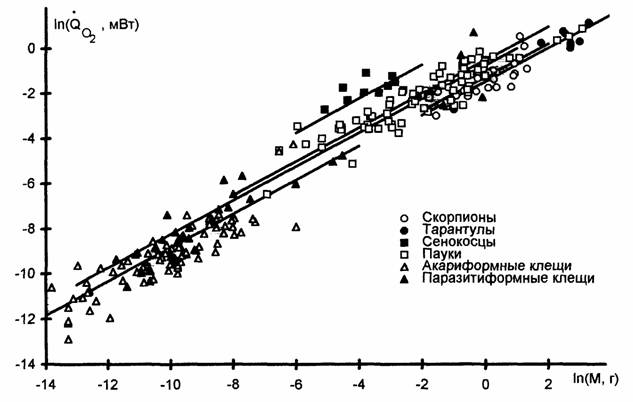
|
| Рис.16. Зависимость скорости потребления кислорода при 20°С от массы тела в классе паукообразных (Зотин и др., 1999). |
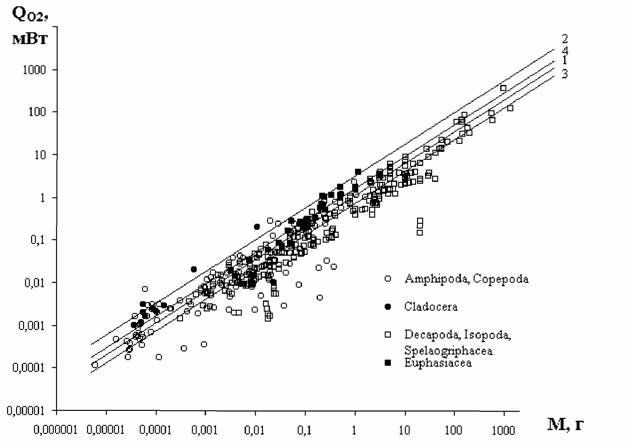
|
| Рис.17. Зависимость скорости потребления кислорода при 20°С от массы тела в классе ракообразных (Алексеева, 1999). |

|
| Рис.18. Зависимость скорости потребления кислорода при 20°С от массы тела в классе насекомых (Алексеева, Зотин, 1996). (1) - пчелы; (2) - муравьи; (3) - бабочки; (4) - клопы. |
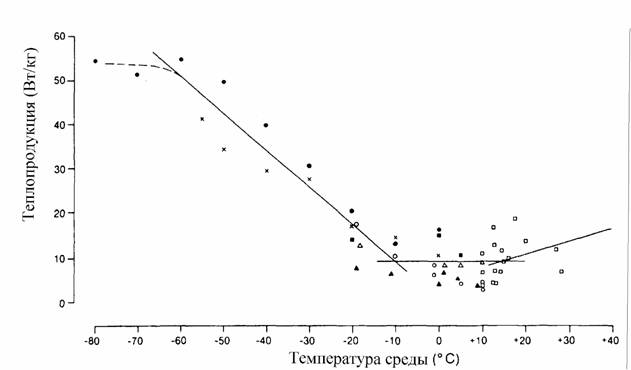
|
| Рис.19. Зависимость удельной теплопродукции роем пчел от температуры среды (Southwick, 1988). |
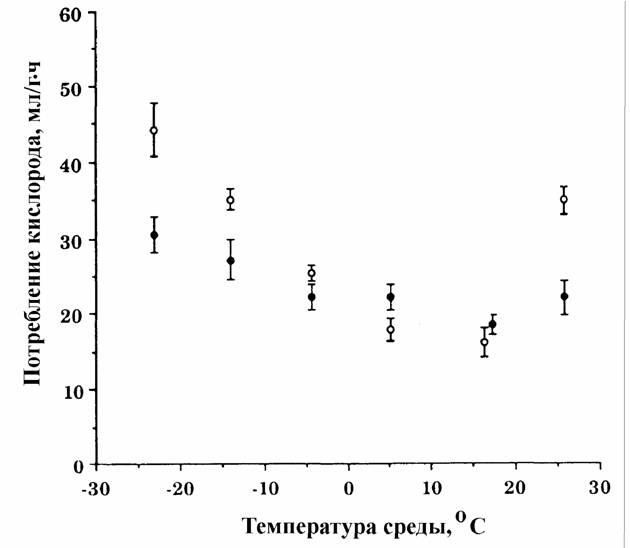
|
| Рис. 20. Интенсивность потребления кислорода группой из 300 пчел (Apis mellifera carnica) летом (незакрашенные кружки) и зимой (закрашенные кружки) (Southwick, 1990). |
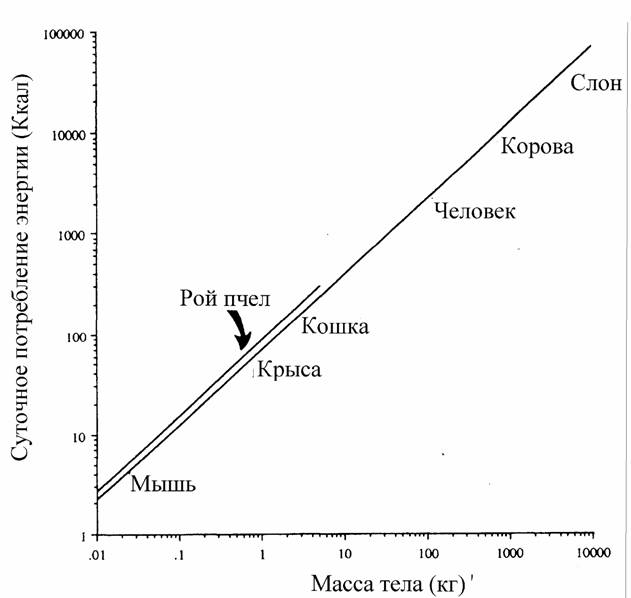
|
| Рис.21. Сравнение дневного потребления энергии роями пчел различной массы при 15°С и плацентарными млекопитающими в термонейтральной зоне (Moritz, Southwick, 1992). |
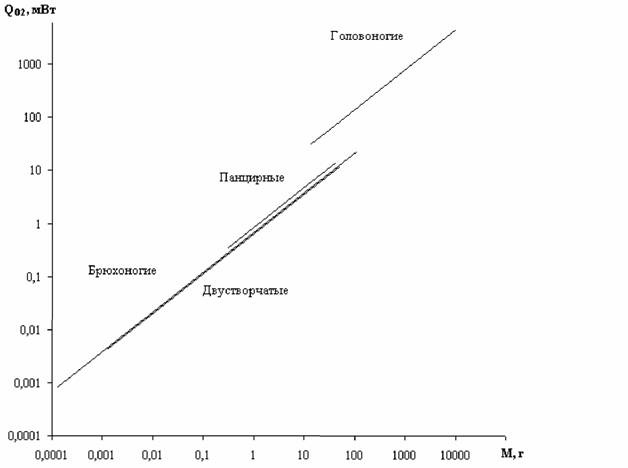
|
| Рис.22. Зависимость скорости потребления кислорода при 20°С от массы тела в типе моллюсков: панцирные, двустворчатые и головоногие моллюски (настоящее сообщение), брюхоногие моллюски (Владимирова, 1999). |
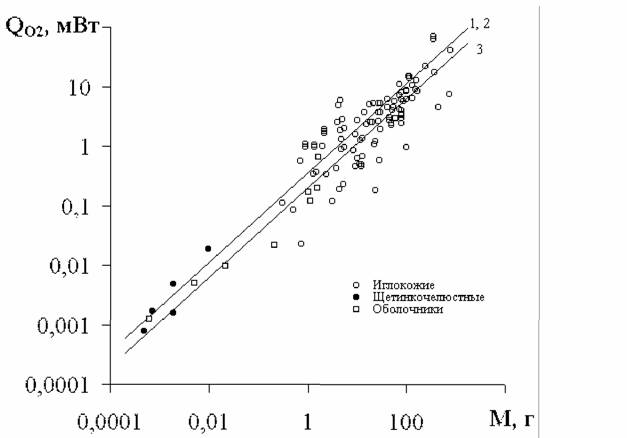
|
| Рис. 23. Зависимость скорости потребления кислорода при 20°С от массы тела у иглокожих (1), щетинкочелюстных (2) и оболочников (3). |
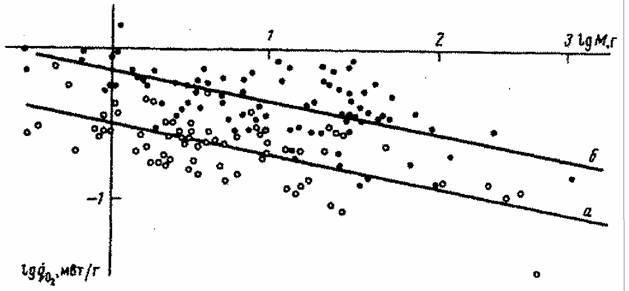
|
| Рис.24. Зависимость интенсивности потребления кислорода при 20°С от массы тела у представителей хвостатых (а) и бесхвостых (б) земноводных (Владимирова, Зотин, 1994). |
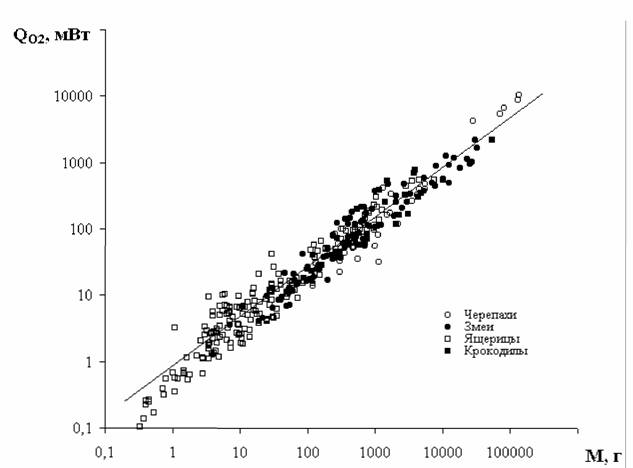
|
| Рис.25. Зависимость скорости потребления кислорода при 20°С от массы тела у представителей разных отрядов и подотрядов класса пресмыкающихся. |
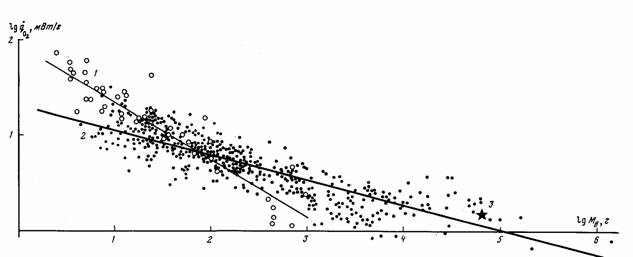
|
| Рис. 26. Зависимость интенсивности потребления кислорода (в термонейтральной зоне) от массы тела в классе млекопитающих: 1 - насекомоядные, 2 - остальные млекопитающие, 3 - человек (Зотин и др., 1990). |
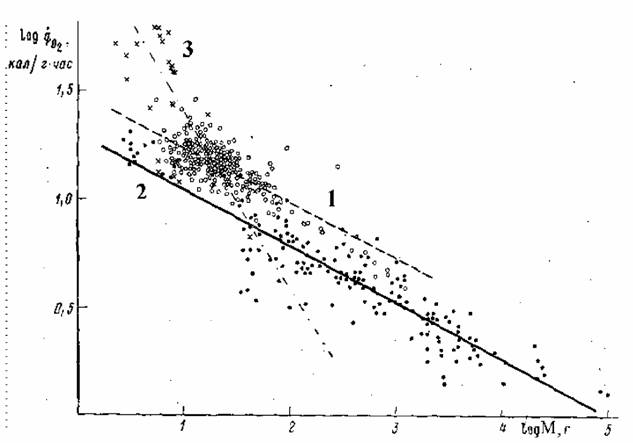
|
| Рис.27. Зависимость интенсивности потребления кислорода (в термонейтральной зоне) от массы тела у воробьиных (1), неворобьиных (2) (Зотин, Криволуцкий, 1982) и колибриобразных (Krüger, Prinzinger, 1982) птиц. |
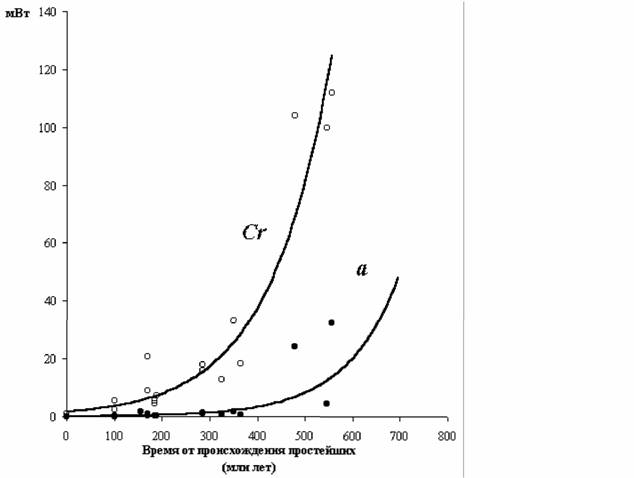
|
| Рис.28. Изменение сопоставимого стандартного обмена (коэффициента а, мВт) и критерия упорядоченности (Cr) в процессе биологической эволюции в разных группах животных (по данным, показанным в табл.41 и 42). |
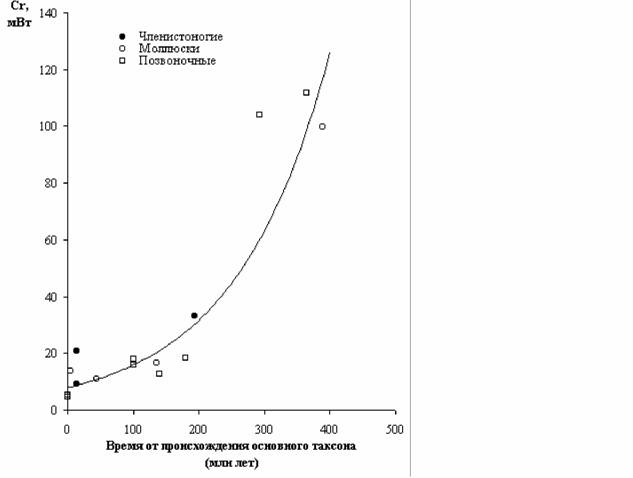
|
| Рис.29. Сравнение темпов прогрессивной эволюции по критерию упорядоченности в типах членистоногих, моллюсков и позвоночных. Время происхождения основного таксона - время возникновения наиболее древних представителей типа. |
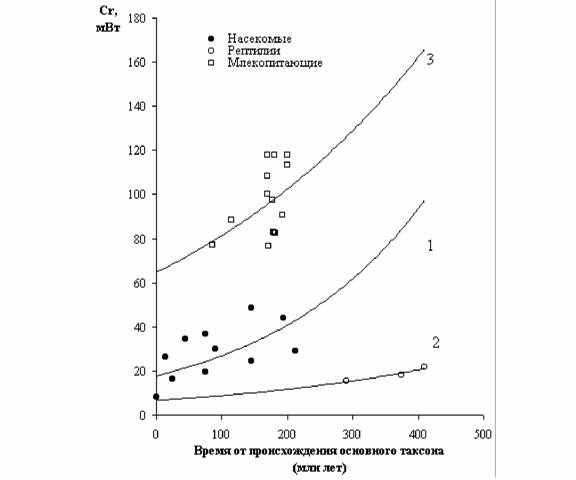
|
| Рис.30. Сравнение темпов прогрессивной эволюции по критерию упорядоченности в классах насекомых (1), брюхоногих моллюсков (2) и млекопитающих (3). Время происхождения основного таксона - время возникновения наиболее древних представителей класса. |
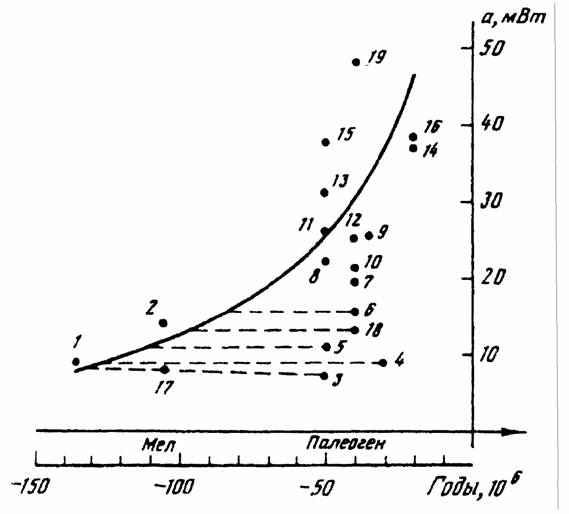
|
| Рис.31. Биоэнергетический прогресс и регресс (изменение коэффициента а, мВт) в процессе эволюции млекопитающих (Зотин и др., 1990). Обозначения: 1 - однопроходные, 2 - сумчатые, 3 - неполнозубые, 4 - ящеры, 5 - сирены, 6 - доманы, 7 - рукокрылые, 8 - зайцеобразные, 9 - приматы, 10 - грызуны, 11 - хищные, 12 - парнокопытные, 13 - ластоногие, 14 - непарнокопытные, 15 - китообразные, 16 - тенреки, 17 - ежи, 18 - землеройки. |
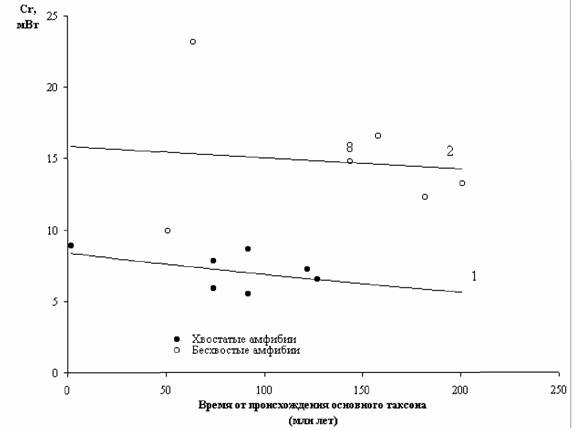
|
| Рис.32. Биоэнергетический регресс (уменьшение коэффициента а, мВт) в процессе эволюции амфибий. Время происхождения основного таксона - время возникновения наиболее древних представителей класса амфибий. |
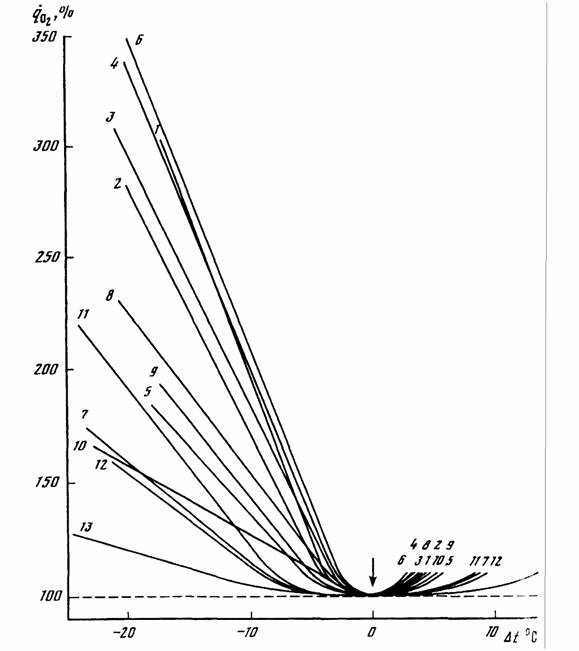
|
| Рис.33. Кривые терморегуляции в разных отрядах класса млекопитающих (Владимирова, Зотин, 1989). Потребление кислорода в % от критической точки (рис.10), где интенсивность дыхания принята за 100% 1 - ехидны; 2 - сумчатые; 3 - насекомоядные; 4 - рукокрылые; 5 - приматы; 6 - неполнозубые; 7 - зайцеобразные; 8 - грызуны; 9 - хищные; 10 - ластоногие; 11 - даманы; 12 - парнокопытные; 13 - непарнокопытные. Стрелкой показано положение критической точки. |
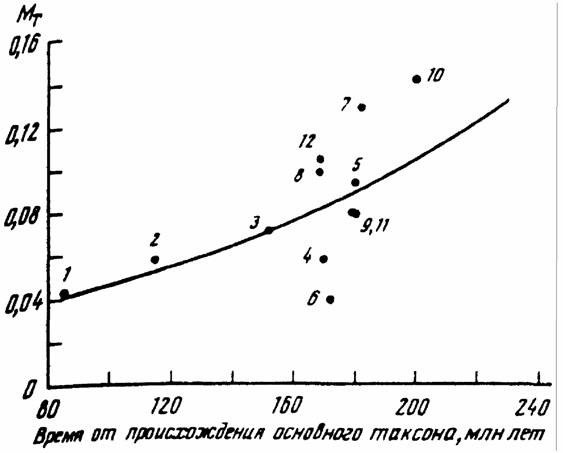
|
| Рис.34. Изменение меры надежности терморегуляции Мт в процессе эволюции млекопитающих. Обозначения те же, что и на рис.33. |

|
| Рис.35. Изменение сопоставимой термонейтральной зоны Z0 в процессе эволюции млекопитающих. Обозначения те же, что и на рис.33. |
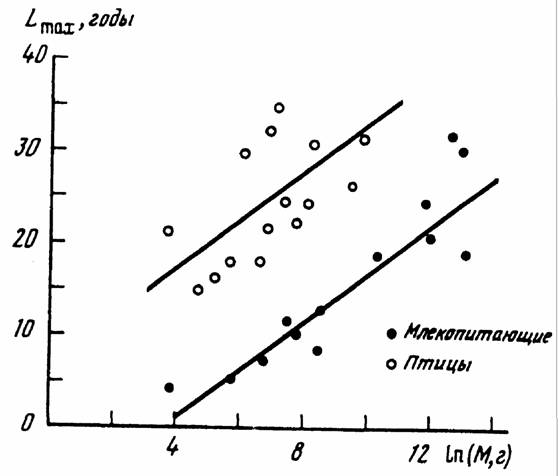
|
| Рис.36. Зависимость максимальной продолжительности жизни животных (Lmax, годы) от массы тела (М, г) у млекопитающих и птиц. |

|
| Рис.37. Зависимость коэффициента энцефализации (сbr) от максимальной продолжительности жизни животных (Lmax, годы) у млекопитающих (Зотин, 19936). |
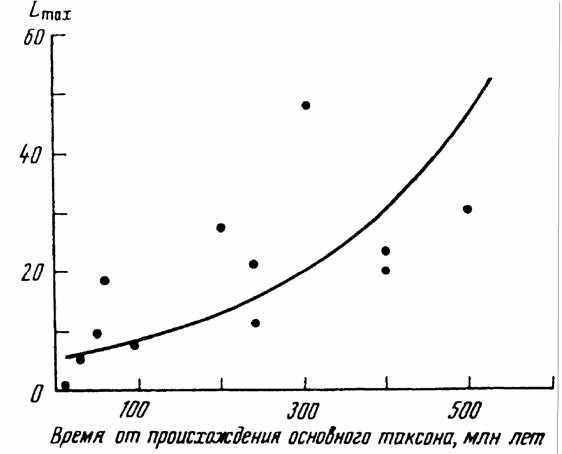
|
| Рис.38. Изменение максимальной продолжительности жизни животных (Lmax, годы) в процессе эволюции (Мирозян, 1989). Время происхождения основного таксона - время возникновения простейших. |
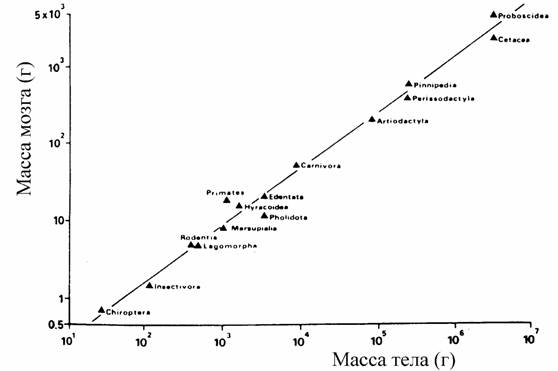
|
| Рис.39. Зависимость массы мозга (Мbг, г) от массы тела (М, г) в классе млекопитающих (Harvey, Bennett, 1983). |
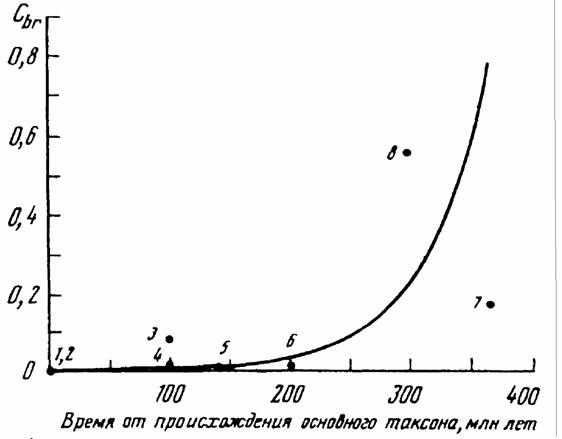
|
| Рис.40. Изменение величины коэффициента энцефализации (сbr) в процессе эволюции позвоночных животных. Обозначения: 1 - Миксины, 2 - Миноги, 3 -Хрящевые рыбы, 4 - Костные рыбы, 5 - Земноводные, 6 - Пресмыкающиеся, 7 - Птицы, 8 - Млекопитающие. Время происхождения основного таксона - время возникновения наиболее древних представителей позвоночных. |
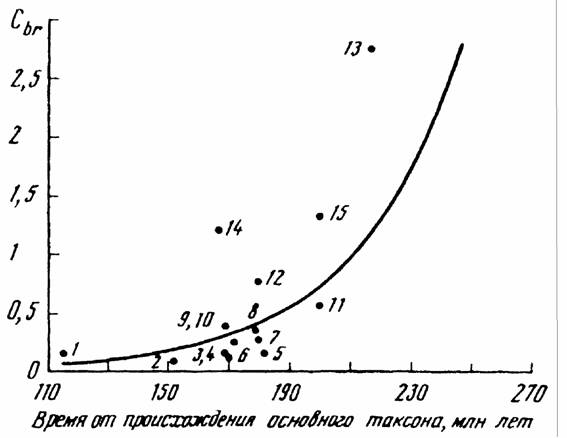
|
| Рис.41. Изменение величины коэффициента энцефализации (сbr) в процессе эволюции млекопитающих. Обозначения: 1 - Сумчатые, 2 - Насекомоядные, 3 - Рукокрылые, 4 - Зайцеобразные, 5 - Грызуны, 6 - Сирены, 7 - Неполнозубые, 8 - Доманы, 9 - Хищные, 10 - Парнокопытные, 11 - Непарнокопытные, 12 - Мозоленогие, 13 - Ластоногие, 14 - Приматы, 15 - Хоботные. Время происхождения основного таксона - время возникновения наиболее древних представителей позвоночных. |
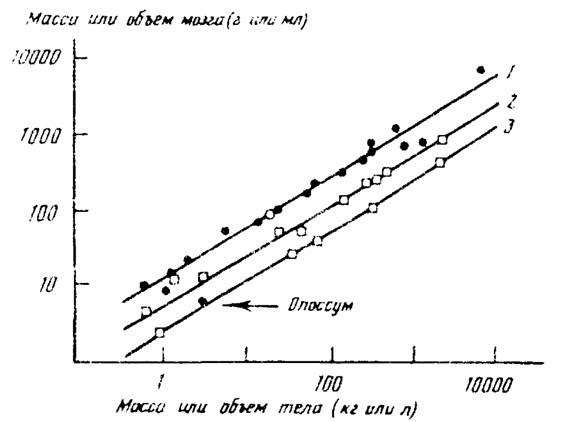
|
| Рис.42. Соотношение между массой мозга (Мbr, г) и массой тела (М, г) у млекопитающих в разные геологические периоды: 1) современное время, 2) олигоцен, 3) эоцен (Jerison,1961). |
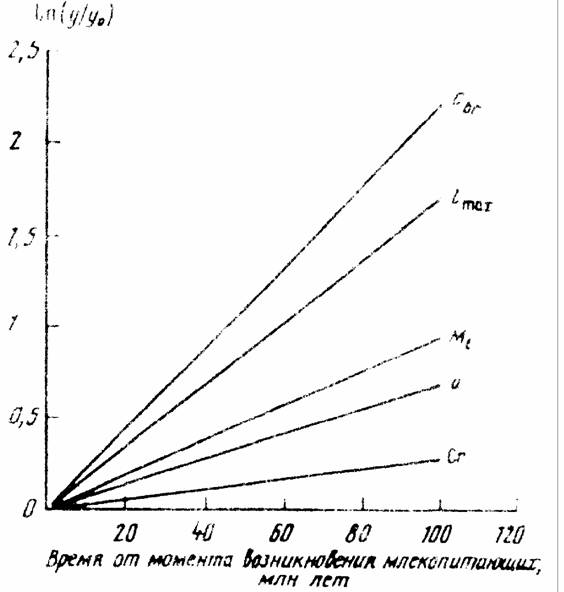
|
| Рис.43. Эволюционный прогресс млекопитающих по критерию упорядоченности (Сr) по сопоставимому стандартному обмену (а), по критерию надежности терморегуляции (Mt), по коэффициенту энцефализации (сbr) и по сопоставимой максимальной продолжительности жизни (lm). По оси ординат - логарифм от соотношения параметра в соответствующий момент времени к величине этого параметра в момент возникновения млекопитающих. |
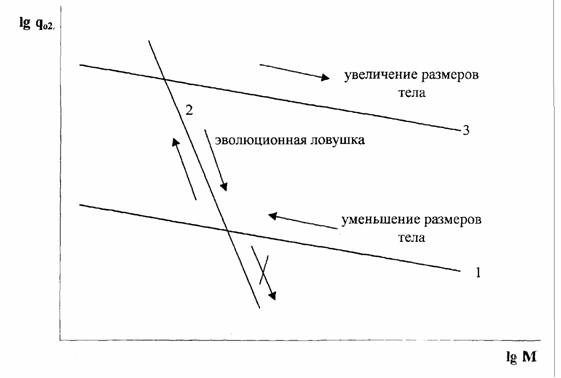
|
| Рис.44. Схема механизма прогрессивной эволюции в результате уменьшения размеров животных. Обозначения: 1 - уменьшение массы тела, 2 - эволюционная ловушка; 3 - прогрессивная эволюция. |
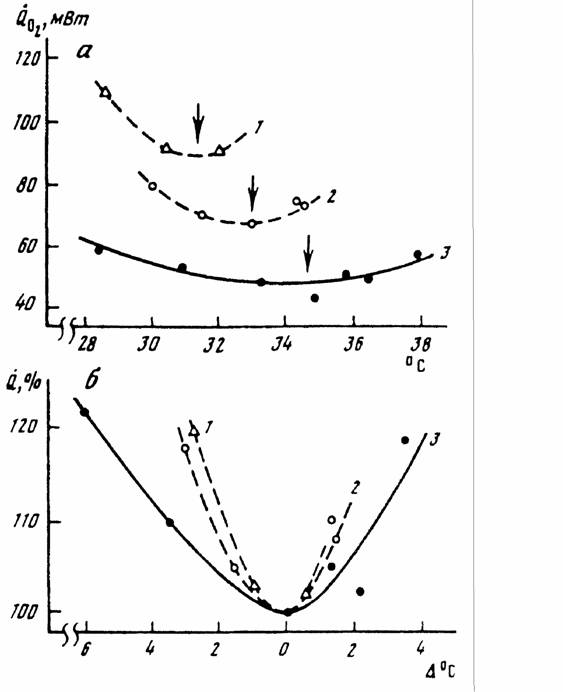
|
Рис.45. Скорость потребления кислорода в мВт ( ) и в % ( ) и в % ( )от
потребления кислорода в критической точке, принятого за 100%, у зеленушки Chloris Moris при акклимации птиц к 2-8°С (1), 15-20°С (2) и 29-32°С (3) (Слоним,1982а). Стрелками показано положение критических точек. )от
потребления кислорода в критической точке, принятого за 100%, у зеленушки Chloris Moris при акклимации птиц к 2-8°С (1), 15-20°С (2) и 29-32°С (3) (Слоним,1982а). Стрелками показано положение критических точек.
|
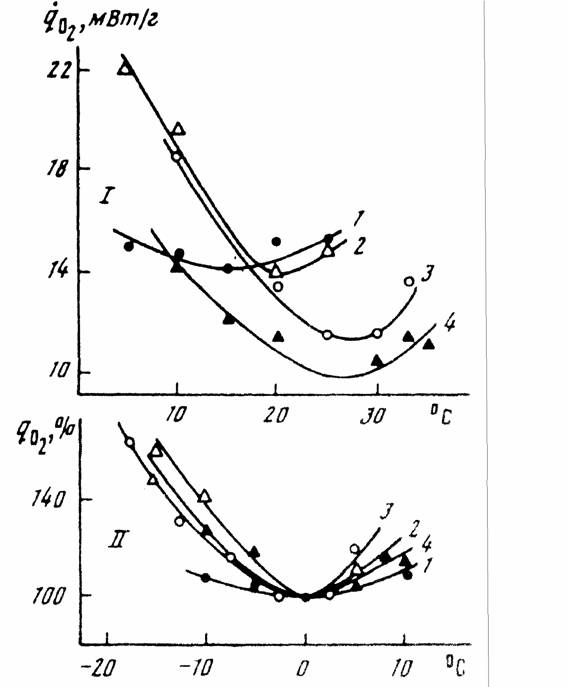
|
| Рис.46. Широтная акклиматизация серой крысы Rattus norvégiens (Руттенбург, 1953). Обозначения: I - Интенсивность потребления кислорода в мВт/г, II - в % от потребления в критической точке, принятого за 100%, 1 - крысы из Мурманска, 2 - из Ленинграда, 3 - из Севастополя, 4 - из Сухуми. |
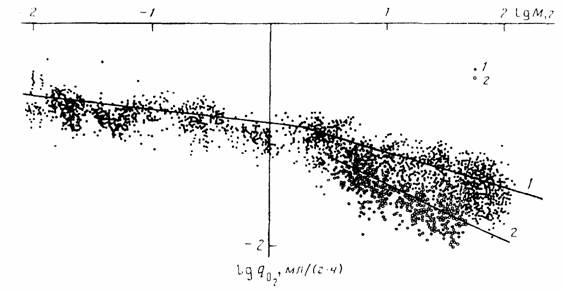
|
| Рис.47. Зависимость интенсивности потребления кислорода при 20°С от массы тела у аксолотлей (1) и амбистом (2) (Владимирова и др., 1993). |
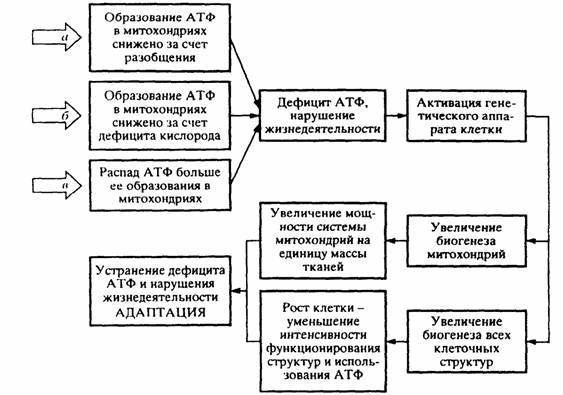
|
| Рис.48. Схема механизмов адаптации организмов к длительному действию холода (а), гипоксии (б) и интенсивных физических нагрузок (в) (Меерсон,1973). |
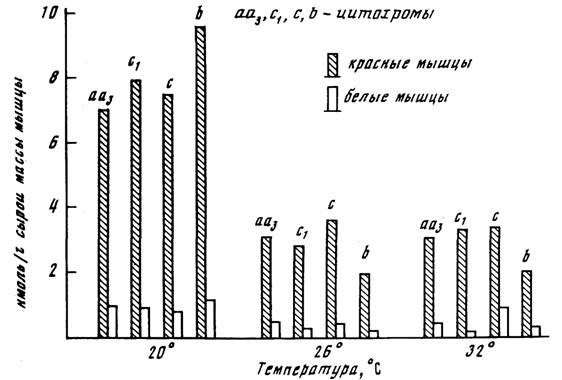
|
| Рис.49. Концентрация цитохромов в скелетных мышцах рыбы Tilapia mossambica, при акклимации к разным температурам (Демин и др., 1989). |
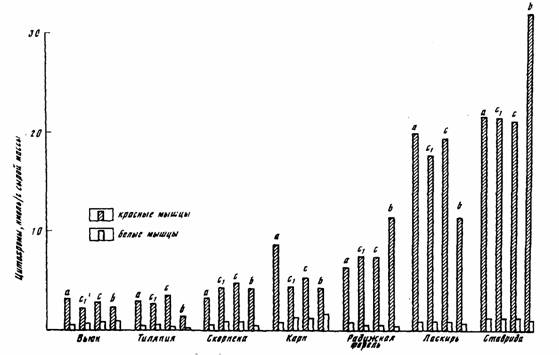
|
| Рис.50. Содержание цитохромов в скелетных мышцах рыб, отличающихся скоростью плаванья (Демин и др. ,1989). |
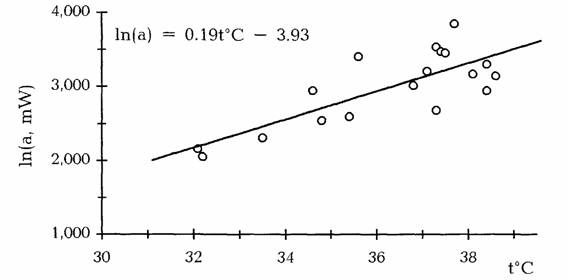
|
| Рис. 51. Зависимость средней температуры тела от сопоставимого стандартного обмена (коэффициента а) у представителей разных отрядов млекопитающих (Zotin et al., 1999). |
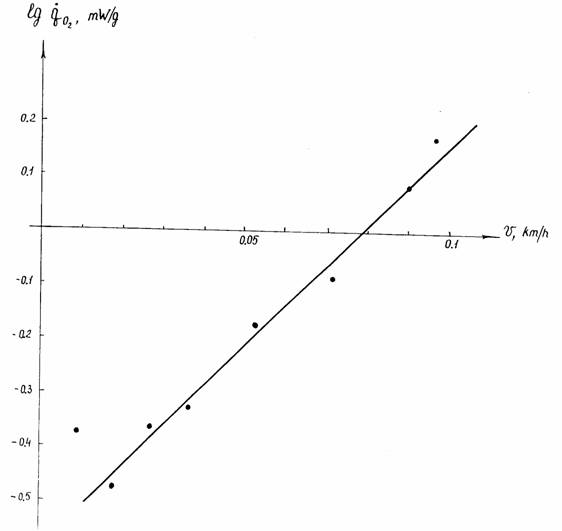
|
| Рис.52. Зависимость активного обмена солнечной рыбы Lepomis gibbosus от скорости плаванья при 20°С (Brett, Sutherland, 1965). |
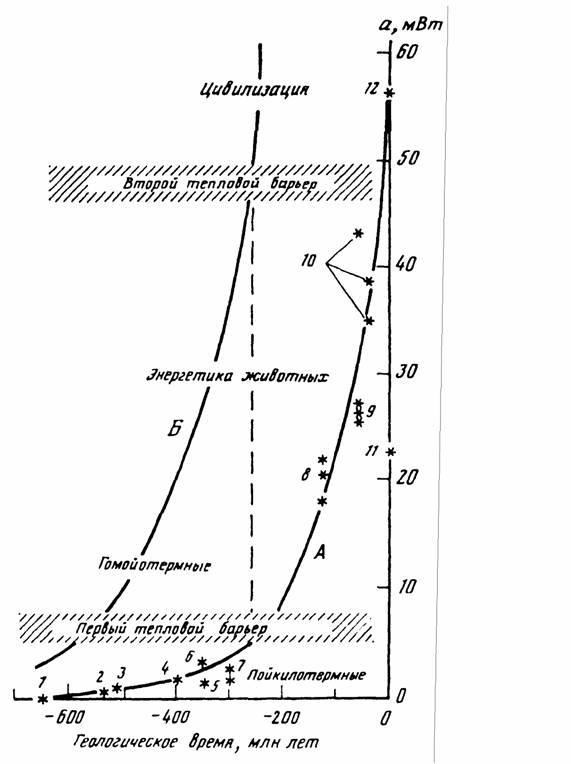
|
| Рис.53. Кривая биоэнергетического прогресса (А), кривая максимального обмена (Б), первый и второй тепловые барьеры. Обозначения такие же, как на рис.6. |
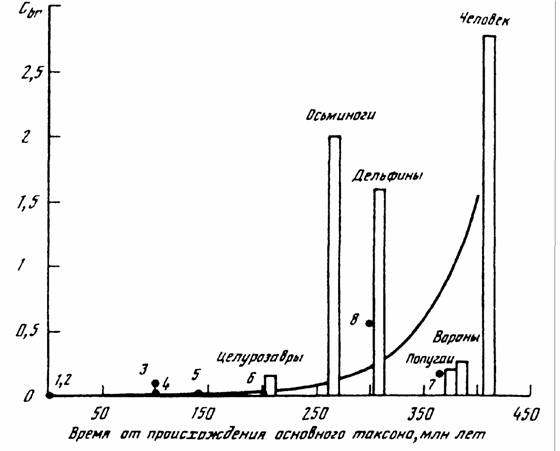
|
| Рис.54. Величина коэффициента энцефализации у целурозавров, осьминогов, дельфинов, попугаев, ворон и человека в сравнении с кривой изменения коэффициента энцефализации в процессе эволюции позвоночных животных. |

|
| Рис.55. Претенденты на создание цивилизаций в прошлом. Динозавры из группы целурозовров. |

|
| Рис.56. Претенденты на создание цивилизации в будущем. Дельфины (рис. Н.В.Илюченко по: Крушинская, Лисицына, 1983). |

|
| Рис.57. Претенденты на создание цивилизаций в будущем. Осминоги (из Kirby, Kirby, 1907). |

|
| Рис.58. Претенденты на создание цивилизаций в будущем. Врановые птицы (рис. Н.В.Илюченко по: Корнишъ, 1907). |
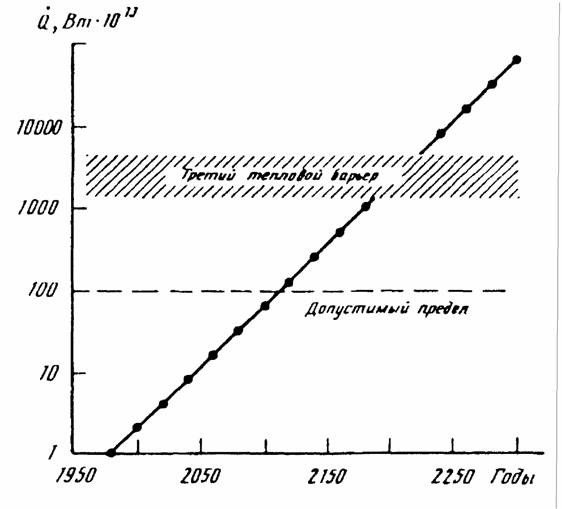
|
| Рис.59. Предполагаемое увеличение энергопроизводства человечеством в ближайшем будущем и третий тепловой барьер. |
ЭЛЕКТРОННОЕ ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ............................................................................................................................. 3
Часть 1. ТЕРМОДИНАМИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИИ....................... 7
Глава 1. Необходимые сведения из термодинамики неравновесных процессов.................. 8
Глава 2. Необходимые сведения из теории биологической эволюции................................. 29
1. Микроэволюция......................................................................................................................................................... 30
Аксиоматика................................................................................................................................................................. 31
Синтетическая теория эволюции............................................................................................................................ 34
2. Макроэволюция......................................................................................................................................................... 36
Биологический и морфо-физиологический прогресс......................................................................................... 36
3. Прогрессивная эволюция....................................................................................................................................... 40
Биоэнергетический прогресс.................................................................................................................................... 42
Совершенствование регуляции и управления..................................................................................................... 49
4. Мегаэволюция............................................................................................................................................................ 52
Механизмы мегаэволюции....................................................................................................................................... 53
Регрессивная эволюция.............................................................................................................................................. 56
Глава 3. Термодинамика и биология........................................................................................ 59
1. Энтропия и негэнтропия........................................................................................................................................ 61
Энтропия........................................................................................................................................................................ 61
Негэнтропия.................................................................................................................................................................. 63
2. Организованные системы...................................................................................................................................... 64
Живые системы............................................................................................................................................................ 68
3. Термодинамические подходы к проблемам биологической эволюции.............................................. 70
Термодинамика и направленность эволюции..................................................................................................... 71
4. Общие соотношения биологической термодинамики............................................................................... 78
Часть 2. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: УСИЛЕНИЕ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА...................................................................................................................................... 82
Глава 1. Критерии биоэнергетического прогресса и прогрессивной эволюции................. 83
1. Стандартный обмен.................................................................................................................................................. 84
Значение массы тела.................................................................................................................................................. 85
Влияние температуры................................................................................................................................................ 88
Акклимация и акклиматизация................................................................................................................................ 92
2. Критерий упорядоченности................................................................................................................................... 93
3. Максимальный обмен........................................................................................................................................... 103
4. Константа Рубнера................................................................................................................................................. 105
Глава 2. Стандартный обмен и критерий упорядоченности у беспозвоночных животных 108
1. Простейшие............................................................................................................................................................... 110
2. Низшие беспозвоночные...................................................................................................................................... 115
3. Черви - Plathelminthes (Плоские черви)........................................................................................................... 122
4. Высшие беспозвоночные...................................................................................................................................... 126
Класс Insecta (Насекомые)........................................................................................................................................ 138
Низшие насекомые - Apterygota............................................................................................................................ 141
Высшие насекомые - Pterygota.............................................................................................................................. 144
Насекомые с неполным превращением.............................................................................................................. 144
Насекомые с полным превращением................................................................................................................... 151
Суперорганизмы........................................................................................................................................................ 158
Макросистематика насекомых............................................................................................................................. 160
Тип Mollusca (Моллюски)....................................................................................................................................... 164
Тип Echinodermata (Иглокожие)........................................................................................................................... 178
Тип Chaetogliatha (Щетинкочелюстные)............................................................................................................ 184
Тип Tunicata (Оболочники).................................................................................................................................... 185
Глава 3. Стандартный обмен и критерий упорядоченности у позвоночных животных. 187
Класс Amphioxi (Ланцетники)............................................................................................................................... 187
Рыбы................................................................................................................................................................................. 188
Класс Amphibia (Земноводные).............................................................................................................................. 197
Класс Reptilia (Пресмыкающиеся)........................................................................................................................ 201
Класс Mammalia (Млекопитающие)..................................................................................................................... 205
Класс Aves (Птицы)..................................................................................................................................................... 212
Глава 4. Прогрессивная эволюция и биоэнергетический прогресс.................................... 218
1. Изменение энергетического обмена в ходе эволюции.............................................................................. 219
2. Биологический и физический смысл эволюционного прогресса........................................................ 233
3. Направление и скорость прогрессивной эволюции и биоэнергетического прогресса................ 238
4. Направление и скорость эволюционного прогресса в соответствии с константой Рубнера.... 245
5. Аллогенез и катагенез............................................................................................................................................ 248
Часть 3. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: СОВЕРШЕНСТВОВАНИЕ РЕГУЛЯЦИИ И УПРАВЛЕНИЯ....................................................................................................... 252
Критерии организованности..................................................................................................... 254
Глава 1. Температурный гомеостаз......................................................................................... 261
1. Совершенствование терморегуляции в процессе прогрессивной эволюции.................................. 263
2. Адаптация к условиям существования.......................................................................................................... 265
3. Скорость совершенствования терморегуляции у млекопитающих в процессе эволюции........ 268
4. Суперорганизмы...................................................................................................................................................... 270
Глава 2. Максимальная продолжительность жизни............................................................. 272
Эволюция максимальной продолжительности жизни животных............................................................... 275
Эволюция максимальной продолжительности жизни в отдельных классах позвоночных животных. 275
Глава 3. Относительные размеры и энергетика мозга......................................................... 278
1. Зависимость массы мозга от массы тела........................................................................................................ 279
2. Зависимость массы мозга от потребления кислорода.............................................................................. 284
3. Изменение организованности животных в процессе эволюции........................................................... 286
Часть 4. МАКРОСКОПИЧЕСКИЕ МЕХАНИЗМЫ БИОЭНЕРГЕТИЧЕСКОГО ПРОГРЕССА.......................................................................................................... 292
Глава 1. Уменьшение размеров тела....................................................................................... 293
Глава 2. Генотипические адаптации....................................................................................... 299
1. Адаптация к низким температурам среды.................................................................................................... 300
2. Адаптация к высокогорью.................................................................................................................................. 308
3. Адаптация к физическим нагрузкам.............................................................................................................. 311
Глава 3. Неотения...................................................................................................................... 313
Глава 4. Митохондриальные механизмы прогрессивной эволюции.................................. 316
Часть 5. БИОЭНЕРГЕТИЧЕСКИЙ ПРОГРЕСС И ТЕПЛОВЫЕ БАРЬЕРЫ 322
Глава 1. Первый тепловой барьер и возникновение терморегуляции............................... 325
Способны ли крокодилы и динозавры к терморегуляции?........................................................................... 329
Глава 2. Второй тепловой барьер и возникновение цивилизаций...................................... 332
1. Увеличение относительных размеров мозга................................................................................................ 335
2. Виртуальные цивилизации................................................................................................................................. 339
3. Социальная деятельность животных.............................................................................................................. 340
Глава 3. Третий тепловой барьер и выход человечества в космическое пространство... 342
1. Ноосфера как следствие принципа наискорейшего спуска.................................................................. 344
2. Принцип наискорейшего спуска, как основа эволюции Вселенной................................................. 347
3. Третий тепловой барьер....................................................................................................................................... 348
ЗАКЛЮЧЕНИЕ...................................................................................................... 351
ЛИТЕРАТУРА............................................................................................................................. 355
РИСУНКИ (1-29):....................................................................................................................... 433
ЭЛЕКТРОННОЕ ОГЛАВЛЕНИЕ............................................................................................. 491