Определение поля - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2 Развитие Половых Признаков. Формирование Половых Признаков У Человека ...
Развитие половых признаков. Формирование половых признаков у человека – один из аспектов его развития, изученный относительно хорошо. Эта область представляет собой пример удачного сочетания теории, наблюдений непосредственно на человеке и экспериментов на животных. Теория подсказывала исследователям направления экспериментов, которые в свою очередь позволяли усовершенствовать саму теорию. Наблюдения генетических аномалий определения пола у человека тоже способствовали развитию теории и, кроме того, указывали на необходимость новых экспериментов.
Принято различать четыре уровня половой дифференцировки:
1) хромосомное определение пола (46 XX или 46 XY);
2) определение пола на уровне гонад (яичники или семенники);
3) фенотипическое определение пола (женский или мужской; внешние половые признаки);
4) психологическое определение пола.
Четвертый уровень будет обсуждаться в разд. 8.2.2.3, первый частично уже затрагивался в разд. 2.2.3. Как отмечалось, анализ аутосомных аберраций, несмотря на его несомненную важность для понимания механизмов нормального развития и его нарушений, до сих пор не позволил выяснить ничего конкретного о механизмах развития. С другой стороны, анализ нарушения числа и структуры половых хромосом дал очень много ценной информации не только о хромосомном определении пола (1-й уровень), но также об определении пола на уровне гонад и фенотипа (уровни 2 и 3, см. ниже и [1068а]). Зачатки гонад у ранних эмбрионов (до 5-й или 6-й недели) не различаются у разных полов и не содержат клеток зародышевого пути. Первичные клетки зародышевого пути у человека можно обнаружить на 3-й неделе эмбрионального развития в эктодерме желточного мешка. Затем под влиянием хемотаксических сигналов они мигрируют в гонады. Эта миграция не зависит от пола; в соответствующих экспериментальных системах женские половые клетки мигрируют в мужские гонады и наоборот. Зачатки гонад могут развиваться в яичники или семенники (уровень 2). В норме направление развития определяется наличием Y-хромосомы; мужские гонады развиваются, если имеется одна Y-хромосома, независимо от числа Х-хромосом. Согласно современным данным, это развитие зависит от H-Y-антигена.
Y-хромосома и H-Y-антиген. Эйчвальд и Силмсер в 1955 г. описали у мыши трансплантационный антиген, контролируемый Y-хромосомой (разд. 3.1.4) [643]. Лишь значительно позднее стало понятным его значение для определения пола и особенно для дифференцировки мужских гонад [1248; 1249; 1341; 1343; 1342]. В настоящее время H-Y-антиген, определяющий пол, обнаружен у нескольких видов. По-видимому, его секретируют мужские первичные клетки зародышевого пути. Как только эти клетки попадают в зачатки гонад, начинается дифференцировка семенников. Этот вывод в значительной степени основан на изучении случаев отклонений в развитии пола, например у фримартин (т.е. коров генетически женского пола, у которых гонады превратились в семенники в результате того, что некоторое количество клеток близнеца мужского пола транспортировалось через общую кровеносную систему в гонады женского эмбриона). В настоящее время получены экспериментальные данные в пользу того, что H-Y-рецепторы имеются на поверхности клеток гонад обоих типов.
Совместная инкубация дезинтегрированной ткани семенников с H-Y-антигеном приводит к сборке структур, напоминающих семенники. Такая сборка может быть индуцирована и в случае клеток женских гонад. Если активность H-Y-антигена подавить добавлением анти-Н-Y-антисыворотки, то возникают структуры, характерные для яичников. Данные, полученные в последнее время для редких случаев в определении пола (например, для мужчин с генотипом XX), удается объяснить, предположив, что H-Y-антиген кодируется не Y-хромосомой, а структурным аутосомным геном, который находится под контролем Y-хромосомы. Этот ген может быть репрессирован у всех индивидов, не имеющих Y-хромосомы; его экспрессия может быть индуцирована факторами, которые в норме определяются Y-хромосомой. Следователь-
138 4. Действие генов
но, возможны конститутивные мутации, при которых будет синтезироваться H-Y-антиген, приводя к развитию мужского фенотипа даже в тех случаях, когда все клетки организма лишены Y-хромосомы. Возможно, однако, что произошла транслокация, в результате которой ген H-Y-антигена переместился с Y-хромосомы на аутосому. С другой стороны, H-Y-антиген обнаруживали (хотя и в низком титре) даже у женщин с синдромом Тернера (ХО), а также у XX-мужчин. Эти и другие наблюдения привели к выводу, что определение пола на уровне гонад может быть пороговым феноменом; для превращения зачатка гонад в семенники необходима некоторая минимальная концентрация H-Y-антигена.
В последнее время роль H-Y-антигена в формировании семенников вызывает некоторые сомнения, которые связаны с новыми наблюдениями на животных и человеке. Например, H-Y-антигены, обусловливающие реакцию отторжения трансплантата и вызывающие цитотоксический эффект, вероятно, не идентичны, поскольку у некоторых животных эти два теста дают противоположные результаты [1114]. Следует учесть, однако, что тест на цитотоксичность связан с практическими трудностями и несколько субъективен [1038].
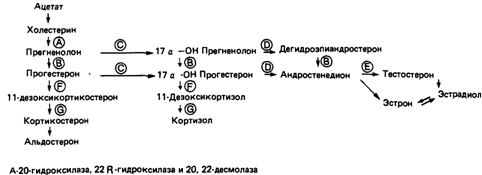
Развитие вторичных половых признаков. Развитие половых признаков обусловлено дифференцировкой гонад. Половые органы формируются из мюллеровых и вольфовых протоков, которые происходят из первичной почки. У женщин мюллеровы протоки развиваются в фаллопиевы трубы и матку, а вольфовы протоки атрофируются. У мужчин вольфовы протоки развиваются в семенные протоки и семенные пузырьки. Под влиянием хорионического гонадотропина матери клетки Лейдига в эмбриональных семенниках синтезируют стероидные гормоны тестостерон и 5-дигидротестостерон. В клетках Сертоли синтезируется гормон, который называют мюллеровым ингибирующим фактором (MIF). Эти гормоны действуют на бипотентные зачатки внешних и внутренних половых органов, прежде всего на вольфовы протоки, мюллеровы протоки и мочеполовой синус. Нормальные индивиды мужского пола развиваются, только если все эти элементы функционируют в нужное время и в надлежащем месте. При их полном отсутствии формируются женские половые признаки; таким образом, развитие женских половых признаков не требует специальных регуляторных факторов, оно в этом смысле является «конститутивным». Незначительные отклонения в работе этой системы на различных уровнях вызывают неполное развитие мужского фенотипа в организме с мужским генотипом (мужской псевдогермафродитизм); анализ таких аномалий позволил получить обширную информацию о нормальной физиологии развития пола. Джост писал по этому поводу: «Становление мужского организма – это длительное, нелегкое и рискованное предприятие, своего рода борьба против имманентного стремления к женственности». Известно по меньшей мере 19 различных дефектов генов, аутосомно-рецессивных или сцепленных с Х-хромосомой, которые вызывают нарушения дифференцировки внешних и внутренних мужских половых признаков. Например, нарушенными могут быть и синтез андрогенов и синтез хорионического гонадотропина, могут отсутствовать рецепторы для этого гормона на клетках Лейдига; дефектными могут оказаться и пять ферментов, участвующих в синтезе тестостерона. Причиной аномалий может быть и нечувствительность клеток вольфовых протоков или мочеполового синуса к тестостерону или 5-дигидротестостерону вследствие дефектов рецепторов.
Пути биосинтеза тестостерона и соответствующие генетические блоки приведены на рис. 4.73. В левой части рисунка (G, F) показаны нарушения, характерные для адреногенитальных синдромов у женщин, но не для псевдогермафродитизма у мужчин. Нарушения, обозначенные А (наследственная липоидная гиперплазия надпочечников), изучены мало; такие индивиды с мужским генотипом не только имеют женские наружные половые органы, но также страдают от сильных нарушений солевого обмена. Это справедливо и для блока В. В правой части рисунка показаны нарушения, которые вызывают псевдогермафро-
4. Действие генов 139
|

| Рис. 4.73. Пути биосинтеза тестостерона и эстрадиола и их генетические дефекты [1068а].
|
дитизм различной степени, не сопровождающийся другими проявлениями адреногенитального синдрома.
Особого внимания заслуживает аутосомно-рецессивный признак - дефект 5-α-редуктазы (не показан на рис. 4.73). В норме этот фермент восстанавливает тестостерон до 5-α-дигидротестостерона в клетках мочеполового синуса. При его отсутствии развиваются нормальные мужские внутренние половые органы (семенные пузырьки, простата), а также вторичные мужские половые признаки, включая развитие мускулатуры, характер волосяного покрова и т.д., за исключением внешних половых органов, которые при поверхностном обследовании можно принять за женские. Отсюда название псевдовагинальная перинеоскротальная гипоспадия (26460).
Эти дефекты ферментов встречаются редко. Существуют более частые синдромы, при которых андрогены не изменены, но соответствующие ткани-мишени полностью или частично невосприимчивы к ним.
Синдром тестикулярной феминизации (31370) [1114]. При рождении эта аномалия никак не проявляется; больные выглядят как обычные девочки и в детском возрасте аномалию обычно удается идентифицировать, только если при паховых грыжах обнаруживаются семенники. Больные с таким синдромом имеют мужской кариотип, и мужские гонады. Термин «тестикулярная феминизация» был предложен Моррисом в 1953 г. [1220]. С наступлением половой зрелости отмечается аменорея, а в большинстве случаев обращает на себя внимание также полное или частичное отсутствие подмышечных волос, волос в области лобка и на теле. У взрослых рост и пропорции типично женские, хотя ноги часто несколько длиннее. Молочные железы хорошо развиты. Пропорции тела таких индивидов соответствуют скорее современным представлениям о женской красоте, чем среднему телосложению, поэтому неудивительно, что больные неоднократно встречались среди манекенщиц.
Влагалище обычно укорочено и заканчивается слепым мешком. Вместо матки часто имеются остатки мюллеровых протоков, а вместо фаллопиевых труб можно найти мышечно-волокнистый тяж. Семенники локализуются в больших половых губах, паховом канале или в брюшной полости, и могут содержать нормальное или даже увеличенное количество клеток Лейдига, продуцирующих гормоны. Сперматогенез обычно отсутствует. Иногда наблюдаются злокачественные опухоли семенников.
У больных андрогены секретируются в нормальных количествах, в особенности тестостерон, и потому можно было бы
140 4 Действие генов
ожидать нормального мужского развития. Наиболее очевидное объяснение механизма этой болезни, которое приводилось в течение многих лет, заключается в ненормальной конечной реакции органов на гормон. Недавно был обнаружен дефект рецепторов андрогенов [1359]. Это подтвердилось в различных экспериментах: например, лечение тестостероном не позволяет добиться изменений в голосе, росте бороды, гипертрофии клитора или (у больных мышей) индукции алкогольдегидрогеназы в почках. Психологическое развитие при тестикулярной феминизации происходит целиком по женскому типу. В настоящее время получены данные, указывающие, что взаимодействие андрогенов и их рецепторов влияет на развитие мозга, электроэнцефалограмму и поведение (разд. 8.2.2.3).
Многие случаи таких отклонений встречаются чисто случайно, однако иногда наблюдаются семейные корреляции, при этом обнаруживается наследование от здоровых женщин и дополнительные случаи среди сестер матерей (матроклинные тетки пробандов). Это позволяло предположить мутацию, сцепленную с Х-хромосомой. Однако, поскольку репродукция у больных полностью нарушена, не удавалось получить и окончательных доказательств сцепления с Х-хромосомой, т.е. наблюдать наследование признака от гемизиготы всеми дочерьми, но не сыновьями. Поэтому нельзя исключить ограниченного полом проявления аутосомно-доминантного гена. Однако, в 1971 г. у мыши была обнаружена мутация с очень близким фенотипом [1025], которая явно была сцеплена с Х-хромосомой. Х-хромосома млекопитающих остается эволюционно стабильной, расположенные в ней гены являются гомологичными у всех до сих пор изученных видов млекопитающих [156]. Более того, рецепторы тестостерона можно обнаружить также в фибробластах, причем гетерозиготы имеют две разные популяции фибробластов - нормальную и неспособную связывать тестостерон, как это предсказывалось гипотезой Лайон [985]. Таким образом, сцепление с Х-хромосомой можно считать установленным. Любопытно, что Паттерсон и Бонньер, основываясь на анализе родословных, еще в 1937 г.

| Рис. 4.74.Пути тестостерона и дигидротестостерона в клетке Различают три типа рецепторных молекул, по-разному седиментирующих в градиенте сахарозы с определенной концентрацией ионов – 4S, 5S и 8S. Попадая в клетку, гормоны связываются с 8S-рецепторами, образуя 8S-гормон-рецепторный комплекс (HRC),последний диссоциирует на 4S-HRCи гормон-неформирующие субъединицы 4S-HRCпревращается в 5S-HRC, который затем транспортируется в ядро Тестикулярная феминизация может возникать в результате различных (полных или частичных) нарушений этой системы [1068а]
|
предполагали (тогда определение пола по генотипу было еще невозможно), что больные являются генотипически мужчинами и признак либо наследуется сцепленно с Х-хромосомой, либо ограничен полом. В результате новых мутаций могут появляться спорадические случаи, давление отбора в этом случае довольно значительное, больные бесплодны, поэтому можно ожидать обнаружения многочисленных новых мутаций (разд. 5.1.3).
Генетическая гетерогенность Полипептидные гормоны, такие как инсулин, связываются с рецепторами, расположенными на мембранах клеток-мишеней (разд 3 8 14) С другой стороны, стероидные гормоны, как например, тестостерон, после проникновения в клетку путем диффузии связываются с цитоплазматическими рецепторами. На рис. 4.74 показан путь тестостерона и дигидротестерона в клетке, начиная со связывания с цитоплазматическим 8S-рецептором, через 4S-рецепторный комплекс к 5S-
4. Действие генов 141
рецепторному комплексу, который транспортируется в ядро. Большинство мутаций, приводящих к тестикулярной феминизации, затрагивают 8S-рецептор. Вполне вероятно, что мутантными могут быть 4S- и 5S-комплексы. Действительно, зарегистрированы случаи тестикулярной феминизации, при которых 8S-рецептор нормален. Кроме того, известны больные с неполной тестикулярной феминизацией, для которых характерно наличие интерсексуальных гениталий. При этом 8S-рецепторы, хотя и обнаруживаются, но в меньшем количестве.
Мы коснулись лишь немногих генетических нарушений определения пола у мужчин. Известно много других, которые согласуются с теоретической концепцией становления пола и благодаря которым мы составили себе представления об этом процессе. Даже у фенотипически нормальных мужчин иногда встречаются генетические дефекты, вызывающие стерильность, например, нарушение формирования из половых клеток жизнеспособных сперматозоидов [1032].
Все темы данного раздела:
Фогель Ф., Мотульски А.
74 Генетика человека: В 3-х т. Т. 2: Пер. с англ.-М.: Мир, 1990. - 378 с, ил.
ISBN 5-03-000286-3
Книга двух известных генетиков из ФРГ и США является фундаментальным учебником
Развитие менделевской парадигмы
Разработка концепций, предложенных Гальтоном и Менделем, приблизила нас к пониманию механизма действия генов. Обсуждение близнецового метода выявило не только его возможности, но и ограничения, свя
Гены и ферменты
4.2.1. Гипотеза «один ген – один фермент»
Первые исследования. После того как в 1902 г. Гэррод указал на связь генетического дефекта при алкаптонурии с неспособност
Гены и ферменты у человека: современный уровень знаний
Круг рассматриваемых вопросов. В каждом случае ферментативного дефекта необходим особый подход в методологии исследований и интерпретации результатов. Ограниченный объем настоящего обзора за
Обнаружение и анализ ферментативных нарушений
Различия в подходах к исследованию человека и нейроспоры. Успехи в изучении ферментативных нарушений у бактерий и нейроспоры были достигнуты благодаря новому направлению исследований. Авторы
Типичные нарушения функций ферментов: ферменты эритроцитов
К настоящему времени подробно изучена группа наследственных заболеваний, связанных с недостаточностью ферментативных систем эритроцитов [933, 1345]. Эритроциты человека – безъядерные клетки, неспос
Мукополисахаридозы
Недостаточность ферментов лизосом. Ферменты или ферментные системы обычно локализуются в одном определенном районе клетки. Например, ферменты системы транспорта электронов и окислительного ф
Выявление гетерозигот
Выявление гетерозигот по ФКУ и гиперфенилаланинемии. Для заболеваний, связанных с нарушением метаболизма (в частности, ФКУ), выявление гетерозигот имеет не только теоретическое, но и практич
Необнаруженные дефекты ферментов
Сколько ферментов у человека и какие дефекты ферментов известны? Некоторые метаболические пути пока еще не выяснены. Поэтому никто не знает точного числа ферментов у человека. Согласно прибл
История изучения гемоглобина
Серповидноклеточная анемия – «молекулярное» заболевание. Изучение гемоглобина человека началось с открытия наследственного заболевания – серповидноклеточной анемии. В 1910 г. Херрик [1121] о
Генетика гемоглобина
Молекулы гемоглобина. Молекула человеческого гемоглобина состоит из четырех полипептидных цепей. Молекула гемоглобина обозначается общей формулой α2β2, котора
Генетика антител и системы антиген/рецептор
Образование антител и их функции. Живые организмы постоянно подвергаются атаке как извне - со стороны бактерий и вирусов, так и изнутри – со стороны клеток, которыев результате случайных соб
Фармакогенетика и экогенетика 4.5.1. Фармакогенетика
Развитие биохимической генетики человека, обнаружение наследственных дефектов ферментов, привели к возникновению новой отрасли генетики - фармакогенетики. Еще Гэррод, основатель биохимической генет
Механизм аутосомной доминантности
Аутосомно-рецессивные заболевания, как правило, обусловлены дефектами ферментов, которые возникают вследствие мутаций в соответствующих генах. Часто удается показать, что фермент имеет аномальную с
Аномальная агрегация субъединиц
Дисфибриногенемии (13480) [1112]. Для этой группы заболеваний характерно, что все симптомы проявляются у гетерозигот. Если такие гетерозиготы несут мутацию в гене, кодирующем белок с субъеди
Аномальные субъединицы нарушают функции мультимерных белков
Гемоглобинопатии. Существует целый ряд клинических форм гемоглобинопатии, которые имеют сходные причины возникновения, связанные с субъединичной структурой гемоглобина. Известно, что в соста
Аномальное ингибирование ферментов по типу обратной связи и структурно аномальные ферменты
Порфирия (17600) [1282; 1217] - понижение ферментативной активности. Установлено, что различные варианты доминантной порфирии (табл. 4.25) являются следствием ферментативных дефектов
Мутации рецепторов
Рецепторы. На поверхности клеточной мембраны имеется множество рецепторов для гормонов, нейромедиаторов и лекарственных препаратов. Должно существовать множество различных мутаций, нарушающи
Наследственные дефекты клеточных мембран
Очевидно, некоторые доминантные нарушения можно объяснить возникновением мутаций, влияющих на мембраны клеток. В качестве примера приведем наследственный сфероцитоз (18290), распространенный тип ге
Доминантно наследуемые опухолевые заболевания
Существуют семьи с доминантно наследуемыми злокачественными и незлокачественными опухолями. Среди них такие часто встречающиеся наследственные заболевания, как нейрофиброматоз, полипоз и ретиноблас
Генетика эмбрионального развития
Результаты исследований по биохимической и молекулярной генетике многое прояснили в структуре генов, а также в генетическом контроле работы ферментов и других белков. В то же время наши знания о ге
Активность генов в раннем развитии
Эмбриональное развитие удобно подразделять на две фазы: раннюю, включающую оплодотворение и несколько первых делений зиготы вплоть до образования гаструлы, и позднюю, на которой закладывается форма
Поздние стадии эмбрионального развития; фенокопии
Формирование систем органов, конечностей, головы и мозга происходит на поздних стадиях эмбрионального развития. Существует ряд типичных наследственных аномалий этого этапа эмбриогенеза, что свидете
Регуляция активности генов у бактерий и эукариот
Отрицательная и положительная регуляция. Многие бактериальные гены активны только тогда, когда их экспрессия необходима, а все остальное время они выключены. Экспрессия таких генов контролир
Эффект дозы генов при трисомиях и картирование генов
Ранние работы по картированию генов с использованием эффекта дозы генов. Обычно при аутосомно-рецессивных аномалиях ферментов их активность у гетерозигот близка к величине, средней для фенот
Другие биохимические аномалии при хромосомных аберрациях
Фетальный и эмбриональный гемоглобины при трисомии 13. Нормальный синтез гемоглобина описан в разд. 4.3.2. ξ, ε и γ-цепи вместе с α-цепями входят в состав ранних э
Изучение хромосомных аберраций на уровне клеток
Фенотипы клеток при хромосомных аберрациях у человека. Биохимические исследования не позволяют сделать определенных выводов о механизмах регуляции эмбриогенеза. Вопрос можно поставить так: ч
Спонтанные мутации
Самое важное свойство генов - их способность передаваться неизменными от поколения к поколению. Однако, если бы генетический материал никогда не менялся, была бы невозможна эволюция. Поскольку суще
Генетические изменения, обусловленные мутациями de novo
Различают следующие типы мутаций:
а) геномные мутации, приводящие к изменению числа хромосом. Геномные мутации часто возникают у растений. При этом может происходить умножение целых
Частота возникновения мутаций (скорость мутирования)
Методы оценки. Чтобы оценить частоту возникновения мутаций, необходимо подсчитать число тех случаев, когда какойлибо признак или наследственная болезнь не обнаруживаются у родителей и других
Нерасхождение хромосом и возраст матери
Статистические данные. Чем старше родители, тем больше вероятность рождения у них ребенка с синдромом Дауна. Этот факт известен уже много лет. На рис. 5.1 представлены частоты новорожденных
У какого пола и в каком из мейотических делений происходит нерасхождение хромосом?
Как было показано в разд. 2.2.1, причина трисомий - нерасхождение хромосом в мейозе. Возникают два вопроса:
5 Мутации 149
Рис. 5.4. Эффект возраста матери в сл
Генные мутации: анализ на фенотипическом уровне
Почти все трисомии, обнаруженные в популяциях человека, возникли в результате мутаций de novo. Больные с этими аномалиями появляются только в отдельных семьях; т. е. здесь мы имеем дело со «споради
Методы оценки частот мутаций
В последующих разделах будут описаны различные методы оценки частот мутаций на основе данных о редких наследственных болезнях. В большинстве случаев эти методы применяются в исследованиях, проводим
Результаты оценки частот мутаций
Оценки, полученные в популяционных исследованиях. Оценки частот мутаций приведены в табл. 5.8. Основной критерий, которым мы руководствовались, включая данные в эту таблицу,-достаточная дост
Частота мутаций и возраст отца
Блестящая идея Вайнберга. В одной из своих работ под названием "Zur Vererbung des Zwergwuchses" (О наследуемой карликовости - нем.) Вайнберг (1912) [1692] обсуждает генетические ос
Возможные различия частот возникновения мутаций у индивидов разного пола
Если большинство мутаций зависит от клеточных делений и репликации ДНК, то следует ожидать не только роста частоты мутаций в мужских половых клетках с увеличением возраста мужчин, но и абсолютно бо
Мутации
детерминирующий дистрофию Дюшенна, находится в Х-хромосоме, кроссинговер (в том числе неравный кроссинговер) может происходить только в женском зародышевом пути.
Косвенн
Герминативноклеточные и соматоклеточные мозаики по доминантным и Х-сцепленным мутациям
Анализ родословных. Если мутация происходит во время раннего развития половых клеток, возможно образование герминативных мозаиков, у которых более или менее значительная часть клеток одной и
Генные мутации: анализ на молекулярном уровне
Анализ нуклеотидных последовательностей ДНК и аминокислотных последовательностей позволяет уточнить представление о механизмах возникновения мутаций. При этом особенно информативными оказались вари
Частоты кодонных мутаций
Какова вероятность мутирования данного кодона в определенном направлении, приводящего к замене одной аминокислоты на другую?
Первая попытка ответить на этот вопрос. Один из авторов
Проблема оценки общей частоты мутаций на геном и на поколение
Условия, необходимые для получения оценки. До сих пор мы рассматривали только частоты мутаций отдельных генов или кодонов. Было бы желательно экстраполировать эти величины и получить общую ч
Мутации в гемоглобиновых генах и генетический код
Аминокислотные замены в полипептидной цепи отражают соответствующие нуклеотидные замены в ДНК. Так как генетический код известен с начала 1960-х годов, изучение аминокислотных замен, особенн
Мутации у микроорганизмов: их вклад в понимание механизма мутаций у человека
Мутации как ошибки репликации ДНК. Данные, полученные на человеке, свидетельствуют о существовании тесной связи между мутациями и клеточными делениями. Импульсом к изучению этой проблемы пос
Изучение генных мутаций в отдельных клетках
В свете успехов генетического анализа микроорганизмов представлялось многообещающим изучение проблем генетики человека на отдельных клетках. Развитие этого подхода описано в разд. 4.2.2.1. Принимая
Соматические мутации
Мутации могут происходить и в половых, и в соматических клетках. Эффект соматической мутации обнаруживается у потомков мутантной клетки, такая мутация делает индивида мозаиком. Мозаик - это особь с
Образование мозаиков по геномным мутациям
Мозаики по геномным мутациям встречаются довольно часто. Сообщалось, например, что в случае синдрома Дауна один мозаик приходится на 48 пациентов, имеющих стандартную трисомию. Исходя из оценки поп
Молекулярные механизмы хромосомной нестабильности и образование опухоли, обусловленное соматической мутацией
Пигментная ксеродерма (27870-27875). Хромосомная нестабильность и существование маркерных хромосом при трех синдромах, с нею сопряженных, наводят на мысль, что повторный разрыв хромосом може
Рак у человека с точки зрения генетики
Разнообразные данные о раке у человека позволяют сделать некоторые обобщения. Менделевское наследование раковых заболеваний человека встречается редко, однако ряд опухолей, особенно доброкачественн
Соматические мутации и старение
Старение и смерть. Люди - единственные живые существа, знающие о неизбежности смерти. Попытки примириться с этой мыслью нашли отражение в культах смерти, имеющих громадное значение в культур
Мутации, индуцированные облучением и химическими мутагенами
Общественный интерес к индуцированным мутациям. В предшествующих разделах говорилось о спонтанных мутациях. Определение «спонтанный» означает, что эти мутации происходят без какой-либо извес
Основные факты и проблемы, поставленные в ходе их анализа
Способность радиации индуцировать мутации. То, что радиация может индуцировать мутации, предполагалось с давних пор, но первые доказательства в пользу этого предположения были получены Мёлле
Проблема оценки генетического риска, обусловленного радиацией и другими мутагенными факторами окружающей среды
Проблема оценки степени риска для человеческой популяции, обусловленного радиацией и всеми другими мутагенными факторами, включает следующие вопросы:
1) Каким образом данный фактор действу
Облучение популяции человека ионизирующей радиацией
Насколько сильно облучается современная человеческая популяция ионизирующей радиацией? Это второй вопрос, на который должен ответить ученый, желающий оценить величину потенциального вреда, причиняе
Насколько может увеличиться частота возникновения спонтанных мутаций9
Как рассчитать возможность увеличения частоты спонтанных мутаций? Это третий вопрос, на который надо ответить, если мы хотим получить оценку возможного генетического повреждения, вызванного радиаци
Данные о соматических хромосомных мутациях, возникающих под воздействием радиации.
Медицинская терапия. Таф и др. (1960) [1659] первыми описали структурные аберрации в хромосомах двух пациентов, облученных при лечении анкилозирующего спондилита. С тех пор выполнено немало
Суть проблемы
История [1384; 1385]. То, что химические вещества индуцируют мутации, предполагалось еще на заре генетики. В своей первой публикации о радиационно-индуцированных мутациях, Мёллер (1927) [156
Исследовательские стратегии при оценке генетического риска, обусловленного химическими мутагенами
На какие вопросы мы должны попытаться получить ответ? Ранее мы упомянули, что перед нами поставлены четыре вопроса: как данный фактор действует на генетический материал; насколько сильно воз
Каким образом химические мутагены действуют на генетический материал?
Планирование тестирующих программ. Из предшествующего обсуждения должно быть ясно, что на первый вопрос - действует ли и как действует определенный фактор на генетический материал - мы не мо
Насколько широким является воздействие агента на человеческую популяцию?
Важный, но часто игнорируемый вопрос. Вопрос о том, насколько широко человеческие популяции подвергаются воздействию данного агента - решающий при получении любой оценки генетической опаснос
Какого увеличения частоты спонтанных мутаций, обусловленного химическими мутагенами, следует ожидать?
Химически индуцированные мутации в сравнении с радиационно-индуцированными мутациями. В разд. 5.2.1.5 мы не могли дать четкого ответа на вопрос об увеличении частоты мутаций под воздействием
Популяционная генетика
Популяционная генетика отвечает на вопросы о том, как реализуются законы Менделя на уровне популяций, как влияют на генетическую структуру популяций такие факторы, как мутационный п
Закон Харди—Вайнберга: генные частоты
Закон Харди—Вайнберга в случае аутосомных генов [124]. Закон Харди—Вайнберга обсуждался в разд. 3.2. Пусть аллели А1 и А2 имеют частоты А1
Генетический полиморфизм
Определение и история вопроса. Полиморфным признаком называется менделевский (моногенный) признак, по которому в популяции присутствуют по крайней мере два фенотипа (и предположительно по кр
Наследственные болезни
Доминантные и. сцепленные с полом рецессивные заболевания. С точки зрения популяционной генетики доминантные и сцепленные с полом рецессивные болезни удобно разделить на две категории.
Систематические изменения генных частот: мутации и отбор
Частоты генов в популяциях остаются неизменными только в отсутствие возмущающих воздействий (см. равновесие Харди— Вайнберга, раздел 3.2). Наиболее важными факторами, изменяющими генные частоты, яв
Математические модели отбора: дарвиновская приспособленность
Область применения математических моделей в теории естественного отбора и их ограничения [124]. Обсуждая отбор, мы будем довольно широко использовать математические модели. В этих моделях де
Отбор, приводящий к изменению генных частот в одном направлении
Используемые символы. Приспособленность генотипа определяется как его способность производить потомство. Она измеряется не в абсолютных, а в относительных единицах, причем приспособленность
Отбор, приводящий к генетическому равновесию
До сих пор мы рассматривали только случаи отбора, приводящего к увеличению частоты одного аллеля за счет уменьшения частоты другого. Рановесие и, следовательно, стабильность генных частот в поколен
Отбор, приводящий к нестабильному равновесию
Отбор против гетерозигот. В популяции может установиться стабильное равновесие, если отбор идет в пользу гетерозигот и против гомозигот. Однако отбор может действовать и в пользу гомозигот п
Другие формулы отбора
Частотно-зависимый отбор [103; 1739]. До сих пор мы считали, что селективные ценности генотипов постоянны. Однако они могут быть функцией частоты генотипов
306 6. Популяционная
История некоторых инфекционных заболеваний.
Какие инфекционные болезни могли влиять в
прошлом на частоту генов в популяциях?
1. Острые инфекционные заболевания, распространявшиеся на целые страны и уносившие значительную ча
Отбор по системе групп крови АВО и другим полиморфным системам
Группы крови АВО и заболеваемость. Ни одна генетическая система у человека не изучена так детально, как система вариантов гемоглобина,
328 6. Популяционная генетика
&n
Отклонение от случайного скрещивания
Впредыдущем обсуждении предполагалось случайное скрещивание и сохранение соотношений Харди—Вайнберга. Однако эти предположения являются абстракцией. В современных аутбредных популя
Кровнородственные браки
6.3.1.1. Коэффициент инбридинга [103]
Все люди – родственники. Родственники определяются как лица, часть генов которых общая по происхождению. Если понимать это опр
Инбридинг, размер изолята и наследственные заболевания
Частота детей с рецессивными и полигенными заболеваниями в кровнородственных и неродственных браках. Пусть аллель, который в гомозиготном состоянии приводит к возникновению рецессивного забо
Практическое применение теории
Предпринималось много попыток оценить реальный генетический груз в популяциях человека. Ранние работы представляли собой практическое приложение теории генетического груза. Однако более поздние исс
Критическая оценка
Теоретическая интерпретация. Все эти результаты, по предложению Мортона [1827], можно интерпретировать в терминах летальных эквивалентов, число которых надо определить. Однако тот факт, что
Более прямые подходы к оценке числа рецессивных генов на индивид
В последнее время используются более прямые подходы к оценке среднего числа рецессивных генов на индивид. Эти исследования ограничиваются генами, приводящими в гомозиготном состоянии к появлению ан
Дифференциация субпопуляций: генетическое расстояние
Реальная структура скрещивания популяций человека. Обычное предположение о том, что в популяциях человека преобладает случайное скрещивание - это абстракция. Выбор брачного партнера вовсе не
Поток генов
Кроме отбора (рассмотренного выше) и случайных флуктуации генных частот (которые будут обсуждаться позднее), на генофонд популяции оказывает большое влияние также поток генов. Для обозначения перен
Генетический дрейф
Детерминистические и стохастические модели. До сих пор наше обсуждение полностью основывалось на менделевских соотношениях и на законе Харди—Вайнберга. Такие популяционно-генетические параме
Генетический дрейф в сочетании с мутационным процессом и отбором
Мутационный процесс. Представим себе большую популяцию, состоящую из множества субпопуляций малого или среднего размера. Распределение аллельных частот q в этих субпопуляциях зависит







Новости и инфо для студентов