Отбор, приводящий к изменению генных частот в одном направлении
Отбор, приводящий к изменению генных частот в одном направлении - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2 Используемые Символы. Приспособленность Генотипа Определяется Как Его ...
Используемые символы. Приспособленность генотипа определяется как его способность производить потомство. Она измеряется не в абсолютных, а в относительных единицах, причем приспособленность оптимального генотипа принимается равной единице. Отклонения от единицы обозначаются через s. Например, если приспособленность генотипа составляет 80% от приспособленности оптимального генотипа (s = 0,2), приспособленность этого генотипа равна 1 -s = 1 — 0,2 = 0,8. Чтобы избежать путаницы со знаками, иногда удобно использовать непосредственную меру приспособленности 1 — s = w. Иногда в литературе s определяется не относительно оптимальной, а относительно средней приспособленности популяции w. Такое обозначение не очень удачно, так как в этом случае приспособленность генотипа изменяется в зависимости от распределения генотипов в популяции. Поэтому мы будем определять приспособленность по отношению к «оптимальному» генотипу. Элиминация гетерозиготного «доминантного» фенотипа. Этот случай простой и встречается часто: какая-либо мутация изменяет фенотип ее носителя так сильно, что его размножение полностью прекращается. К таким последствиям у человека приво-
6. Популяционная генетика 297
дят все нарушения числа хромосом, большинство хромосомных аберраций и многие доминантные генные мутации (разд. 5.1.3). Хромосомные аберрации обнаруживаются прямым методом, однако вновь возникающие доминантные мутации можно выявить только в том случае, если они передаются потомству. Следовательно, доминантные мутации, приводящие к потере репродуктивной способности, невозможно обнаружить. О ее присутствии можно судить только на основании обследования тех немногочисленных индивидов, которые все же дали потомство, или по эффекту отцовского возраста. Возможно, что некоторые болезни и врожденные аномалии вызываются такими до сих пор не выявленными доминантными мутациями.
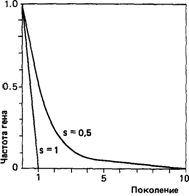
Частичная элиминация аутосомных доминантных мутаций. Большинство вызываемых доминантными мутациями заболеваний уменьшает среднюю репродуктивную способность их носителей. В отсутствие противодействующих факторов, таких как мутационный процесс, эта потеря генов приводит в каждом поколении к уменьшению генных частот на некоторую величину, которая зависит от интенсивности отбора против носителей мутации (рис. 6.8). Пусть р- частота доминантного аллеля A, a q — частота рецессивного нормального аллеля а. Пусть отбор идет с достаточно высокой интенсивностью, а скорость мутирования μ
Рис. 6.8. Уменьшение частоты доминантного гена в отсутствие вновь возникающих мутаций при s = 1 и s = 0,5; s - коэффициент отбора.
низка (разд. 5.3.1). Тогда гомозиготами АА можно пренебречь, поскольку их частота р2 очень низка. Частоты генотипов равны:
Элиминация особей Аа из популяции на поколение равна приблизительно 2ps. Поскольку только половина их генов - А, утрата генов А на поколение составляет 1/2(2ps)= ps. Генетическое равновесие поддерживается, если эта утрата компенсируется возникновением мутаций:
ps = μ(l -p)≈μ.
Отсюда равновесная частота р
= μ/s. (6.1)
Интересны два крайних случая:
1) Гетерозиготы не размножаются, т. е. s=1, w21= 0. В соответствии с выводом, сделанным нами в предыдущем разделе, частота доминантного аллеля р будет равна скорости мутирования μ.
2) Ген не является селективно невыгодным (т.е. s = 0, w21 = 1). При таких условиях равновесие не возникает; частота доминантного аллеля монотонно возрастает: μ/s ®∞
3) наконец, рассмотрим промежуточный случай, когда доминантный ген испытывает некоторое давление направленного против него отбора, например s= 1/3, w21 = 2/3; конечная частота доминантного гена будет равна утроенной скорости мутирования.
Против большинства патологий действует отбор определенной интенсивности. Их частота в популяции определяется равновесием между мутационным процессом и отбором. Конечно, определяющим фактором здесь является скорость мутирования. Частота заболевания растет до тех пор, пока число пораженных индивидов не уве-
298 6. Популяционная генетика
дичится настолько, что интенсивность направленного против них отбора уравновесит скорость мутирования и будет достигнуто генетическое равновесие. Это равновесие стабильно. Если число пораженных превысит равновесную величину, утрата неблагоприятного аллеля в результате отбора будет происходить быстрее его возникновения путем мутирования, и в следующем поколении его частота уменьшится. С другой стороны, если число больных станет ниже равновесной величины, мутационный процесс приведет к появлению большего числа неблагоприятных генов, чем то количество, которое отметается естественным отбором, и частота гена будет увеличиваться, пока вновь не установится равновесие.
Ослабление отбора. Уравнение 6.1 можно использовать для определения последствий релаксации отбора. Предположим, что в результате медицинского вмешательства некоторые фенотипические проявления доминантной мутации были устранены, что привело к снижению селективной невыгодности аллеля с s1= 1/2 до s2= ¼ . Пусть – равновесная частота для s2. Тогда из уравнения (6.1)
Следовательно, новая частота равна удвоенной старой. Новое равновесие будет достигнуто через несколько поколений.
Ослабление отбора против ретинобластомы. Ретинобластома — это злокачественная опухоль глаза, поражающая детей раннего возраста. Подавляющее большинство случаев возникновения ретинобластомы в популяциях являются спорадическими. Однако заболевание часто встречается в семьях и имеет аутосомное доминантное наследование с 90%-ной пенетрантностью. Все билатеральные, но только 10-12% унилатеральных спорадических случаев обусловлены вновь возникающими мутациями.
В прежнее время ретинобластома почти всегда приводила к фатальному исходу: больные погибали в детском возрасте. В 1865 г. ван Графе впервые применил энуклеацию пораженного глаза; позднее в практику были введены дополнительные терапевтические меры - рентгеновское облучение и коагуляция светом; в настоящее время около 90% больных с унилатеральным и 80% больных с билатеральным поражениями могут вылечиться и иметь детей [1887]. Из всех вновь возникающих мутаций, определяющих наследственную форму ретинобластомы, 68% вызывают билатеральный и 32% унилатеральный тип поражения. Изменение частоты этого заболевания можно прогнозировать следующим образом.
Путь Хо-частота больных наследственной регинобластомой в популяции; поскольку это заболевание очень редкое, частотой гомозигот можно пренебречь, а частоту нормального аллеля принять за q к 1; тогда Хопочти равна частоте гетерозигот (2pq s 2p). Пусть перед релаксацией отбора st= 1. После появления эффективных методов лечения давление отбора в случае унилатеральной формы стало sv = 0,1 (только 10% больных унилатеральной формой ретинобластомы умирают в детском возрасте; 90% из них выживают и имеют потомство); давление отбора в случае билатеральной формы sB= 0,2 (80% больных выживают и имеют потомство). Отсюда получаем следующую суммарную оценку s2(сохранившийся после релаксации отбор против ретинобластомы):
Новый коэффициент отбора после ослабления его интенсивности равен 16,8% от прежнего (до релаксации) коэффициента отбора (s=l). Частота гетерозигот в (п + 1)-м поколении, Хп+vполучается из частоты гетерозигот в n-м поколении, Хп, путем применения следующей приближенной формулы:
Из этой рекуррентной формулы можно получить общую формулу для Хn.
Таким образом, новая равновесная частота гетерозигот X представляет собой сумму членов геометрической прогрессии с начальным членом Хо = 2μ и знаменателем (1 — s2):
Этот результат может быть получен также из уравнения (6.1). Отсюда следует, что через несколько поколений после применения эффективного лечения наследственная ретинобластома станет встречаться примерно в 6 раз чаще. Используя реальные оценки, полученные для современных популяций (см. табл. 5.8), получаем Х0= 2μ= 1,2 х 10–5:
6. Популяционная генетика 299
Из этой величины можно вычислить суммарную частоту всех случаев ретинобластомы, включая ненаследственные случаи, если известна доля наследственных случаев среди всех случаев ретинобластомы в популяции перед ослаблением отбора. Общая частота ретинобластомы приблизительно равна 4 х 10–5; следовательно, около 2,8 х 10–5 – частота наследственной формы. Отсюда получаем оценку равновесной частоты после ослабления отбора:
7,14 х 10–5 + 2,8 х 10–5 = 9,94 х 10–5.
Это значит, что частота ретинобластомы возросла с ≈1:25000 до 1:10000, т.е. на 150%. Кроме того, до ослабления отбора наследственными являлись около 30% случаев, тогда как при новом состоянии равновесия наследственными окажутся уже ≈ 72%. Это равновесие устанавливается относительно быстро (рис. 6.9): через 9 поколений частота наследственной ретинобластомы увеличилась более чем в 4 раза по сравнению с соответствующей частотой до ослабления отбора (при равновесии X = 5,95 Х0). На рис. 6.9 приведены также расчеты для двух альтернативных случаев: s2= 0,4 и s2= 0, т. е. отбор отсутствует. В последнем случае частота возрастает линейно и равновесие не устанавливается. Можно все же надеяться, что теоретически рассчитанные частоты на станут фактическими, естественный отбор будет частично заменен искусственным, благодаря генетическому консультированию и добровольному контролю над рождаемостью, осуществляемому носителями данной мутации.
Отбор путем полной элиминации гомозигот. Гомозиготы по многим рецессивным аутосомным заболеваниям обычно не размножаются. Рассмотрим опять два аллеля А и а, имеющие частоты р и q. Однако пусть теперь против гомозигот аа действует отбор:
s=1; w22= 0.
Частота зигот указана в таблице. В общем случае частоты генов в двух последовательных поколениях п и n + 1 связаны следующим образом:
(6.2)
Эта формула выражает рекуррентное соотношение между последовательными частичными суммами (суммами первых п
Рис. 6.9. Ожидаемое увеличение частоты ретинобластомы в популяции, обусловленное ослаблением отбора. Ордината: 1 = частота наследственной ретинобластомы (– – = 2 х скорость мутирования) при таком отборе против ретинобластомы, когда погибают все гомозиготы. Абсцисса: число поколений. Три предположения относительно коэффициента отбора s: отбор отсутствует (s = 0), слабое давление отбора (s = = 0,168); более сильное давление отбора (s — 0,4) [1887].
членов) гармонической последовательности1). Из нее можно получить общую формулу для генной частоты qnчерез п поколений:
(6.3)
Изменение частоты гена за поколение равно
(6.4)
Из уравнения (6.3) следует, что qn= q0/2, если nq0= 1. Следовательно, через п = 1/q0поколений частота гена уменьшится вдвое.
Рассмотрим важное практическое при-
1) Гармоническая последовательность – это такая последовательность, последовательные частичные суммы которой являются величинами, обратными соответствующим членам арифметической последовательности:
В данном конкретном случае b = 1; с = k = и0.
300 6. Популяционная генетика
ложение полученных соотношений. Если размножение гомозигот по кистозному фиброзу будет полностью отсутствовать, как это повлияет на частоту данного гена? Коэффициент отбора против гомозигот s= 1.
АА
Аа
аа
Частота аллеля, а
До отбора
р2
2pq
q2
После отбора
Частота гена, определяющего это заболевание, q0= 0,02, что соответствует частоте гомозигот в панмиктической популяции, равной q02 = 0,0004, т. е. 4 больных на 10000 индивидов. Через п = 1/0,02 = 50 поколений q0уменьшится с 0,02 до 0,01. Принимая длительность поколения за 30 лет, такое уменьшение частоты гена в два раза потребует 1500 лет. В случае более редких заболеваний, таких, как галактоземия, q0 = 0,005, q2= 1:40 000; частота гена уменьшится вдвое за 200 поколений, или 6000 лет.
Таким образом, попытки снизить число рецессивных генов в популяции путем прекращения репродукции гомозигот крайне неэффективны. Более того, в этих расчетах даже не принимались во внимание вновь возникающие мутации.
Частичная элиминация гомозигот. При некоторых рецессивных заболеваниях гомозиготы не теряют репродуктивной способности полностью, однако их биологическая приспособленность сильно понижается. Предположим, что репродуктивная способность гомозигот снизилась на s (1 > s > 0). Частоты генов до и после отбора можно взять из таблицы 6.5. Они дают такую частоту рецессивного аллеля в следующем поколении:
(6.5)
Соотношение между частотами рецессивного аллеля в двух последовательных поколениях задается формулой
Таблица 6.5. Частичная элиминация гомозигот
АА
Аа
аа
Сумма
Перед отбором
Р2
2pq
q2
Приспособленность
1 - s
После отбора
Р2
2pq
q2(1-s)
1 - sq2
Вероятно, эта рекуррентная формула не имеет общего решения. Изменение частоты на поколение равно
Δq зависит как от qn, так и от рп= 1 — qn. Если одна из этих величин мала, Δq также мало. Например, для s = 0,2 q: 0,99 0,50 0,01 Δq: – 0,00244 – 0,0263 – 0,0000198.
При очень малых значениях q, Δq аппроксимируется – sq2.
Некоторые расчеты. Для того чтобы определить изменение q за большое число поколений, Δq можно заменить на
Интегрируя обе части по п поколениям, получаем
Вычисление числа поколений (п), необходимого для того, чтобы q изменилось на определенную величину (при s — 0,01 против гомозигот), дает следующие величины:
Уменьшение q
Число поколений
0,9999-0,9990
0,9990 0,9900
0,9900-0,5000
0,5000-0,0200
5 189
0,0200-0,0100
0,0100-0,0010
90 231
0,0010-0,0001
900 230
6 Популяционная генетика 301
За это время могут произойти события, которые значительно сильнее повлияют на генетический состав популяции.
Гаметический отбор. До сих пор мы предполагали, что отбор действует на стадии зиготы. Однако отбор может происходить уже на стадии гаметы. Из-за своего генетического состава некоторые гаметы имеют меньшую вероятность оплодотворения по сравнению с остальными.
Если какая-нибудь мутация оказывает влияние на вероятность оплодотворения, по этой мутации будет наблюдаться отклонение от нормального расщепления. Предполагается, что именно этим объясняется аномальное расщепление, наблюдающееся при синдроме Алпорта и оптической атрофии Лебера (разд. 3.1.5). Для всех других заболеваний соответствие с менделевским расщеплением обычно считается очевидным. На самом деле это может быть не так, поскольку для демонстрации незначительных отклонений от менделевских соотношений необходимы большие выборки. Однако возникновение существенных отклонений маловероятно.
В случае гамет, несущих сбалансированные и несбалансированные транслокации, нарушение расщепления несомненно имеет место, однако его механизм еще не выяснен (разд. 2.2.2). Если относительный вклад гамет А и а в генофонд следующего поколения составляет 1 и 1 — s, а их частоты равны р и q, то их вклад в генофонд следующего поколения будет р и q(1 — s) соответственно. Тогда изменение генной частоты на поколение
Все приведенные выше примеры на самом деле не очень просты. Поскольку отбор, вероятно, по-разному действует на зародышевые клетки разного пола, ситуация усложняется в случае таких аномалий, как например, транслокации.
С формальной точки зрения, гаметический отбор почти идентичен отбору против гомозигот и гетерозигот с промежуточным значением приспособленности (табл. 6.6).
Таблица 6.6.Коэффициент отбора 2s против гетерозигот и s против гомозигот
Фогель Ф., Мотульски А.
74 Генетика человека: В 3-х т. Т. 2: Пер. с англ.-М.: Мир, 1990. - 378 с, ил.
ISBN 5-03-000286-3
Книга двух известных генетиков из ФРГ и США является фундаментальным учебником
Развитие менделевской парадигмы
Разработка концепций, предложенных Гальтоном и Менделем, приблизила нас к пониманию механизма действия генов. Обсуждение близнецового метода выявило не только его возможности, но и ограничения, свя
Гены и ферменты
4.2.1. Гипотеза «один ген – один фермент»
Первые исследования. После того как в 1902 г. Гэррод указал на связь генетического дефекта при алкаптонурии с неспособност
Гены и ферменты у человека: современный уровень знаний
Круг рассматриваемых вопросов. В каждом случае ферментативного дефекта необходим особый подход в методологии исследований и интерпретации результатов. Ограниченный объем настоящего обзора за
Обнаружение и анализ ферментативных нарушений
Различия в подходах к исследованию человека и нейроспоры. Успехи в изучении ферментативных нарушений у бактерий и нейроспоры были достигнуты благодаря новому направлению исследований. Авторы
Типичные нарушения функций ферментов: ферменты эритроцитов
К настоящему времени подробно изучена группа наследственных заболеваний, связанных с недостаточностью ферментативных систем эритроцитов [933, 1345]. Эритроциты человека – безъядерные клетки, неспос
Мукополисахаридозы
Недостаточность ферментов лизосом. Ферменты или ферментные системы обычно локализуются в одном определенном районе клетки. Например, ферменты системы транспорта электронов и окислительного ф
Выявление гетерозигот
Выявление гетерозигот по ФКУ и гиперфенилаланинемии. Для заболеваний, связанных с нарушением метаболизма (в частности, ФКУ), выявление гетерозигот имеет не только теоретическое, но и практич
Необнаруженные дефекты ферментов
Сколько ферментов у человека и какие дефекты ферментов известны? Некоторые метаболические пути пока еще не выяснены. Поэтому никто не знает точного числа ферментов у человека. Согласно прибл
История изучения гемоглобина
Серповидноклеточная анемия – «молекулярное» заболевание. Изучение гемоглобина человека началось с открытия наследственного заболевания – серповидноклеточной анемии. В 1910 г. Херрик [1121] о
Генетика гемоглобина
Молекулы гемоглобина. Молекула человеческого гемоглобина состоит из четырех полипептидных цепей. Молекула гемоглобина обозначается общей формулой α2β2, котора
Генетика антител и системы антиген/рецептор
Образование антител и их функции. Живые организмы постоянно подвергаются атаке как извне - со стороны бактерий и вирусов, так и изнутри – со стороны клеток, которыев результате случайных соб
Фармакогенетика и экогенетика 4.5.1. Фармакогенетика
Развитие биохимической генетики человека, обнаружение наследственных дефектов ферментов, привели к возникновению новой отрасли генетики - фармакогенетики. Еще Гэррод, основатель биохимической генет
Механизм аутосомной доминантности
Аутосомно-рецессивные заболевания, как правило, обусловлены дефектами ферментов, которые возникают вследствие мутаций в соответствующих генах. Часто удается показать, что фермент имеет аномальную с
Аномальная агрегация субъединиц
Дисфибриногенемии (13480) [1112]. Для этой группы заболеваний характерно, что все симптомы проявляются у гетерозигот. Если такие гетерозиготы несут мутацию в гене, кодирующем белок с субъеди
Аномальные субъединицы нарушают функции мультимерных белков
Гемоглобинопатии. Существует целый ряд клинических форм гемоглобинопатии, которые имеют сходные причины возникновения, связанные с субъединичной структурой гемоглобина. Известно, что в соста
Мутации рецепторов
Рецепторы. На поверхности клеточной мембраны имеется множество рецепторов для гормонов, нейромедиаторов и лекарственных препаратов. Должно существовать множество различных мутаций, нарушающи
Наследственные дефекты клеточных мембран
Очевидно, некоторые доминантные нарушения можно объяснить возникновением мутаций, влияющих на мембраны клеток. В качестве примера приведем наследственный сфероцитоз (18290), распространенный тип ге
Доминантно наследуемые опухолевые заболевания
Существуют семьи с доминантно наследуемыми злокачественными и незлокачественными опухолями. Среди них такие часто встречающиеся наследственные заболевания, как нейрофиброматоз, полипоз и ретиноблас
Генетика эмбрионального развития
Результаты исследований по биохимической и молекулярной генетике многое прояснили в структуре генов, а также в генетическом контроле работы ферментов и других белков. В то же время наши знания о ге
Активность генов в раннем развитии
Эмбриональное развитие удобно подразделять на две фазы: раннюю, включающую оплодотворение и несколько первых делений зиготы вплоть до образования гаструлы, и позднюю, на которой закладывается форма
Поздние стадии эмбрионального развития; фенокопии
Формирование систем органов, конечностей, головы и мозга происходит на поздних стадиях эмбрионального развития. Существует ряд типичных наследственных аномалий этого этапа эмбриогенеза, что свидете
Регуляция активности генов у бактерий и эукариот
Отрицательная и положительная регуляция. Многие бактериальные гены активны только тогда, когда их экспрессия необходима, а все остальное время они выключены. Экспрессия таких генов контролир
Эффект дозы генов при трисомиях и картирование генов
Ранние работы по картированию генов с использованием эффекта дозы генов. Обычно при аутосомно-рецессивных аномалиях ферментов их активность у гетерозигот близка к величине, средней для фенот
Изучение хромосомных аберраций на уровне клеток
Фенотипы клеток при хромосомных аберрациях у человека. Биохимические исследования не позволяют сделать определенных выводов о механизмах регуляции эмбриогенеза. Вопрос можно поставить так: ч
Определение поля
Развитие половых признаков. Формирование половых признаков у человека – один из аспектов его развития, изученный относительно хорошо. Эта область представляет собой пример удачного сочетания
Спонтанные мутации
Самое важное свойство генов - их способность передаваться неизменными от поколения к поколению. Однако, если бы генетический материал никогда не менялся, была бы невозможна эволюция. Поскольку суще
Генетические изменения, обусловленные мутациями de novo
Различают следующие типы мутаций:
а) геномные мутации, приводящие к изменению числа хромосом. Геномные мутации часто возникают у растений. При этом может происходить умножение целых
Частота возникновения мутаций (скорость мутирования)
Методы оценки. Чтобы оценить частоту возникновения мутаций, необходимо подсчитать число тех случаев, когда какойлибо признак или наследственная болезнь не обнаруживаются у родителей и других
Нерасхождение хромосом и возраст матери
Статистические данные. Чем старше родители, тем больше вероятность рождения у них ребенка с синдромом Дауна. Этот факт известен уже много лет. На рис. 5.1 представлены частоты новорожденных
Генные мутации: анализ на фенотипическом уровне
Почти все трисомии, обнаруженные в популяциях человека, возникли в результате мутаций de novo. Больные с этими аномалиями появляются только в отдельных семьях; т. е. здесь мы имеем дело со «споради
Методы оценки частот мутаций
В последующих разделах будут описаны различные методы оценки частот мутаций на основе данных о редких наследственных болезнях. В большинстве случаев эти методы применяются в исследованиях, проводим
Результаты оценки частот мутаций
Оценки, полученные в популяционных исследованиях. Оценки частот мутаций приведены в табл. 5.8. Основной критерий, которым мы руководствовались, включая данные в эту таблицу,-достаточная дост
Частота мутаций и возраст отца
Блестящая идея Вайнберга. В одной из своих работ под названием "Zur Vererbung des Zwergwuchses" (О наследуемой карликовости - нем.) Вайнберг (1912) [1692] обсуждает генетические ос
Мутации
детерминирующий дистрофию Дюшенна, находится в Х-хромосоме, кроссинговер (в том числе неравный кроссинговер) может происходить только в женском зародышевом пути.
Косвенн
Генные мутации: анализ на молекулярном уровне
Анализ нуклеотидных последовательностей ДНК и аминокислотных последовательностей позволяет уточнить представление о механизмах возникновения мутаций. При этом особенно информативными оказались вари
Частоты кодонных мутаций
Какова вероятность мутирования данного кодона в определенном направлении, приводящего к замене одной аминокислоты на другую?
Первая попытка ответить на этот вопрос. Один из авторов
Проблема оценки общей частоты мутаций на геном и на поколение
Условия, необходимые для получения оценки. До сих пор мы рассматривали только частоты мутаций отдельных генов или кодонов. Было бы желательно экстраполировать эти величины и получить общую ч
Мутации в гемоглобиновых генах и генетический код
Аминокислотные замены в полипептидной цепи отражают соответствующие нуклеотидные замены в ДНК. Так как генетический код известен с начала 1960-х годов, изучение аминокислотных замен, особенн
Изучение генных мутаций в отдельных клетках
В свете успехов генетического анализа микроорганизмов представлялось многообещающим изучение проблем генетики человека на отдельных клетках. Развитие этого подхода описано в разд. 4.2.2.1. Принимая
Соматические мутации
Мутации могут происходить и в половых, и в соматических клетках. Эффект соматической мутации обнаруживается у потомков мутантной клетки, такая мутация делает индивида мозаиком. Мозаик - это особь с
Образование мозаиков по геномным мутациям
Мозаики по геномным мутациям встречаются довольно часто. Сообщалось, например, что в случае синдрома Дауна один мозаик приходится на 48 пациентов, имеющих стандартную трисомию. Исходя из оценки поп
Рак у человека с точки зрения генетики
Разнообразные данные о раке у человека позволяют сделать некоторые обобщения. Менделевское наследование раковых заболеваний человека встречается редко, однако ряд опухолей, особенно доброкачественн
Соматические мутации и старение
Старение и смерть. Люди - единственные живые существа, знающие о неизбежности смерти. Попытки примириться с этой мыслью нашли отражение в культах смерти, имеющих громадное значение в культур
Мутации, индуцированные облучением и химическими мутагенами
Общественный интерес к индуцированным мутациям. В предшествующих разделах говорилось о спонтанных мутациях. Определение «спонтанный» означает, что эти мутации происходят без какой-либо извес
Основные факты и проблемы, поставленные в ходе их анализа
Способность радиации индуцировать мутации. То, что радиация может индуцировать мутации, предполагалось с давних пор, но первые доказательства в пользу этого предположения были получены Мёлле
Облучение популяции человека ионизирующей радиацией
Насколько сильно облучается современная человеческая популяция ионизирующей радиацией? Это второй вопрос, на который должен ответить ученый, желающий оценить величину потенциального вреда, причиняе
Суть проблемы
История [1384; 1385]. То, что химические вещества индуцируют мутации, предполагалось еще на заре генетики. В своей первой публикации о радиационно-индуцированных мутациях, Мёллер (1927) [156
Популяционная генетика
Популяционная генетика отвечает на вопросы о том, как реализуются законы Менделя на уровне популяций, как влияют на генетическую структуру популяций такие факторы, как мутационный п
Закон Харди—Вайнберга: генные частоты
Закон Харди—Вайнберга в случае аутосомных генов [124]. Закон Харди—Вайнберга обсуждался в разд. 3.2. Пусть аллели А1 и А2 имеют частоты А1
Генетический полиморфизм
Определение и история вопроса. Полиморфным признаком называется менделевский (моногенный) признак, по которому в популяции присутствуют по крайней мере два фенотипа (и предположительно по кр
Наследственные болезни
Доминантные и. сцепленные с полом рецессивные заболевания. С точки зрения популяционной генетики доминантные и сцепленные с полом рецессивные болезни удобно разделить на две категории.
Систематические изменения генных частот: мутации и отбор
Частоты генов в популяциях остаются неизменными только в отсутствие возмущающих воздействий (см. равновесие Харди— Вайнберга, раздел 3.2). Наиболее важными факторами, изменяющими генные частоты, яв
Математические модели отбора: дарвиновская приспособленность
Область применения математических моделей в теории естественного отбора и их ограничения [124]. Обсуждая отбор, мы будем довольно широко использовать математические модели. В этих моделях де
Отбор, приводящий к генетическому равновесию
До сих пор мы рассматривали только случаи отбора, приводящего к увеличению частоты одного аллеля за счет уменьшения частоты другого. Рановесие и, следовательно, стабильность генных частот в поколен
Отбор, приводящий к нестабильному равновесию
Отбор против гетерозигот. В популяции может установиться стабильное равновесие, если отбор идет в пользу гетерозигот и против гомозигот. Однако отбор может действовать и в пользу гомозигот п
Другие формулы отбора
Частотно-зависимый отбор [103; 1739]. До сих пор мы считали, что селективные ценности генотипов постоянны. Однако они могут быть функцией частоты генотипов
306 6. Популяционная
История некоторых инфекционных заболеваний.
Какие инфекционные болезни могли влиять в
прошлом на частоту генов в популяциях?
1. Острые инфекционные заболевания, распространявшиеся на целые страны и уносившие значительную ча
Отклонение от случайного скрещивания
Впредыдущем обсуждении предполагалось случайное скрещивание и сохранение соотношений Харди—Вайнберга. Однако эти предположения являются абстракцией. В современных аутбредных популя
Кровнородственные браки
6.3.1.1. Коэффициент инбридинга [103]
Все люди – родственники. Родственники определяются как лица, часть генов которых общая по происхождению. Если понимать это опр
Инбридинг, размер изолята и наследственные заболевания
Частота детей с рецессивными и полигенными заболеваниями в кровнородственных и неродственных браках. Пусть аллель, который в гомозиготном состоянии приводит к возникновению рецессивного забо
Практическое применение теории
Предпринималось много попыток оценить реальный генетический груз в популяциях человека. Ранние работы представляли собой практическое приложение теории генетического груза. Однако более поздние исс
Критическая оценка
Теоретическая интерпретация. Все эти результаты, по предложению Мортона [1827], можно интерпретировать в терминах летальных эквивалентов, число которых надо определить. Однако тот факт, что
Более прямые подходы к оценке числа рецессивных генов на индивид
В последнее время используются более прямые подходы к оценке среднего числа рецессивных генов на индивид. Эти исследования ограничиваются генами, приводящими в гомозиготном состоянии к появлению ан
Дифференциация субпопуляций: генетическое расстояние
Реальная структура скрещивания популяций человека. Обычное предположение о том, что в популяциях человека преобладает случайное скрещивание - это абстракция. Выбор брачного партнера вовсе не
Поток генов
Кроме отбора (рассмотренного выше) и случайных флуктуации генных частот (которые будут обсуждаться позднее), на генофонд популяции оказывает большое влияние также поток генов. Для обозначения перен
Генетический дрейф
Детерминистические и стохастические модели. До сих пор наше обсуждение полностью основывалось на менделевских соотношениях и на законе Харди—Вайнберга. Такие популяционно-генетические параме


 = μ/s. (6.1)
= μ/s. (6.1)





 (6.2)
(6.2)
 (6.3)
(6.3) (6.4)
(6.4) В данном конкретном случае b = 1; с = k = и0.
В данном конкретном случае b = 1; с = k = и0.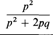


 (6.5)
(6.5)










Новости и инфо для студентов