рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Медицина
- /
- Отбор по системе групп крови АВО и другим полиморфным системам
Реферат Курсовая Конспект
Отбор по системе групп крови АВО и другим полиморфным системам
Отбор по системе групп крови АВО и другим полиморфным системам - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2 Группы Крови Аво И Заболеваемость. Ни Одна Генетическая Система У Чело...
Группы крови АВО и заболеваемость. Ни одна генетическая система у человека не изучена так детально, как система вариантов гемоглобина,
328 6. Популяционная генетика
G6PD и малярии. Однако мы рассмотрим некоторые проблемы, возникающие при анализе таких систем, на примере, гораздо более сложном и противоречивом: группах крови АВО. Как уже отмечалось выше, один аспект отбора по этой системе - серологическая несовместимость матери и плода - общепризнан, хотя относительно интенсивности отбора общего мнения нет. Эта несовместимость приводит к нестабильному равновесию и к медленному изменению частот генов (разд. 6.2.1.4). В отсутствие других форм отбора полиморфизм по АВО должен постепенно исчезнуть. Однако, вопреки этому предсказанию, он присутствует почти во всех популяциях человека. Следовательно, по этой системе должны существовать другие селективные факторы. Имеются ли данные, подтверждающие их наличие?
Группы крови АВО и инфекционные заболевания. В настоящее время продемонстрирована ассоциация групп крови АВО с очень многими заболеваниями (разд. 3.7.2) Например, лица с группой крови А чаще заболевают раком, тогда как лица, имеющие группу крови 0, более подвержены к язве желудка и двенадцатиперстной кишки. Связь с группами крови показана и для ревматизма, иммунный механизм которого бесспорен [211]: риск заболевания ревматизмом самый низкий среди лиц группы 0. Хотя эта ассоциация, возможно, повышает вероятность достижения носителями группы 0 пожилого возраста, она вряд ли оказывает влияние на естественный отбор, поскольку ревматизм, как правило, поражает людей среднего и более старшего возраста, т. е. после завершения репродуктивного периода. Обнаруженные корреляции групп крови системы АВО с заболеваниями демонстрируют фундаментальное влияние антигенов этой системы на физиологию организма. Например, данные по ревматизму свидетельствуют о том, что это влияние может иметь какое-то отношение к иммунному ответу [1789]. Даже ассоциация группы А с раковыми заболеваниями и группы 0 с язвой желудка может быть обусловлена различиями иммунного ответа.
Инфекционные заболевания оказывают специфическое воздействие на иммунный ответ организма. Если группы крови АВО влияют на иммунный ответ, то отбор, возникающий из-за дифференциальной восприимчивости к инфекционным заболеваниям, может привести к дифференциальной детской и юношеской смертности.
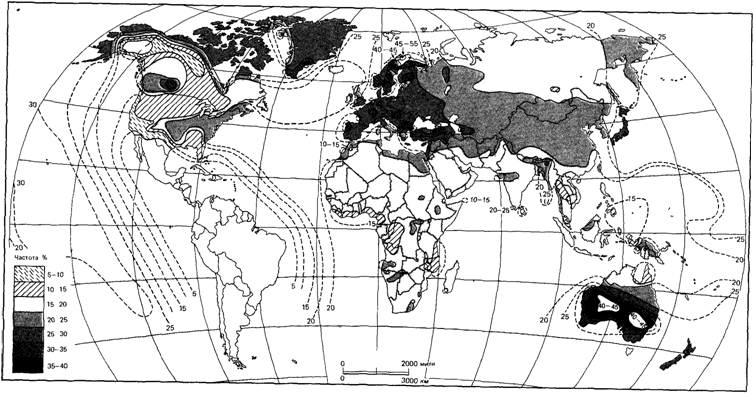
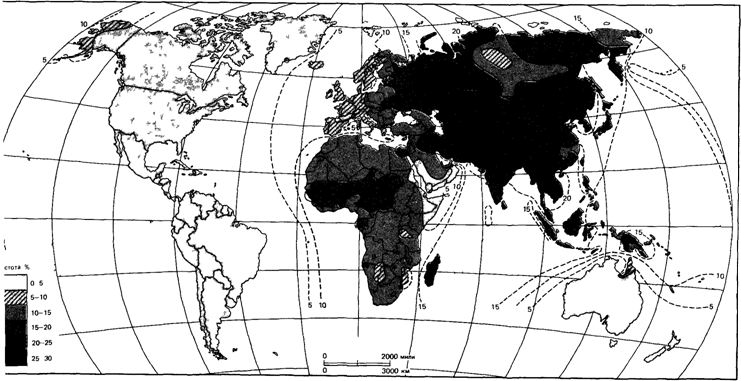
Распространение аллелей АВО в мировом населении. Распределение аллелей А, В и 0 (рис. 6.27-6.29) [144] свидетельствует о наличии естественного отбора по данной системе. Если бы отбора не было, а распределение отражало только случайные колебания генных частот, то в популяции присутствовали бы все комбинации генных частот, возможные в системе из трех аллелей. В действительности дело обстоит по-другому: наблюдается только небольшое число из возможных комбинаций [1732].
Некоторые сведения относительно механизма отбора можно получить, рассмотрев распределение аллеля 0. Обычно этот аллель с высокой частотой встречается в популяциях, которые в течение длительного времени находились в относительной изоляции (например, коренное население Австралии, Полинезии, Арктики и северной Сибири). В Европе, в некоторых изолированных популяциях также наблюдается высокая частота этого аллеля (ирландцы, баски, исландцы, жители Корсики и Сардинии). Особенно высокая частота аллеля 0 отмечается у индейцев Центральной и Южной Америки, что отличает их от других популяций. Различия по частотам других полиморфных систем, например Rh, свидетельствуют против гипотезы о том, что когда-то население этих областей имело высокую частоту аллеля 0. Имеющиеся данные предполагают наличие естественного отбора по этому аллелю. Какая форма отбора могла привести к повышению аллельных частот в сравнительно изолированных популяциях или, наоборот, к их понижению в популяциях, расположенных «в русле» мировых миграций? Одна из вероятных возможностей - влияние инфекционных заболеваний, особенно крупных эпидемий прошлого.
В разделе 6.2.1.6 перечислены следующие группы заболеваний, которые, возможно, имеют селективное значение:
а) эпидемические заболевания, например чума, холера, оспа;
б) хронические болезни, например туберкулез и сифилис;
|
| Рис. 6.27.Распределение частоты аллеля А в популяциях коренного населения разных регионов планеты [144] |
|
| Рис. 6.28. Распределение частоты аллеля В в популяциях коренного населения разных регионов планеты [144] |

|
| Рис. 6.29. Распределение частоты аллеля 0 в популяциях коренного населения разных регионов планеты [144]. |
332 6. Популяционная генетика
в) кишечные инфекции (в основном детские);
г) тропические болезни (в детском и юношеском возрасте).
В отличие от полиморфных вариантов гемоглобина, которые встречаются только в тропических областях, полиморфизм по системе АВО распространен по всему миру. Следовательно, тропические болезни вряд ли играют роль селективного фактора в этой системе. Для формулировки рабочей гипотезы можно использовать три факта.
1. Население Центральной и Южной Америки до открытия этого континента Колумбом было почти полностью изолировано. Возможно, что для американских популяций того времени была характерна особая группа инфекционных заболеваний, отсутствующих в остальной части мирового населения, например сифилис и связанные с ним болезни, вызываемые трепонемой. Частота группы крови 0 в этих популяциях очень высока. Есть ли данные в пользу того, что носители группы крови 0 менее восприимчивы к заражению Treponema pallidum - микроорганизма, вызывающего сифилис?
2. Эпидемии чумы многократно опустошали Европу, в основном поражая густонаселенные области. В краевых и частично изолированных популяциях, которые, вероятно, были затронуты чумой в меньшей степени, группа 0 обычно встречается с высокой частотой. Имеются ли данные, свидетельствующие о большей устойчивости носителей группы 0 к возбудителю чумы?
3. До середины 70-х годов во многих странах регистрировались случаи оспы. Имеются статистические данные о частоте встречаемости и смертности от оспы, особенно для населения Африки и Индии. Для этих регионов известно распределение групп крови АВО. Если носители какой-то группы крови обладают повышенной восприимчивостью к оспе, то в областях с высоким уровнем заболеваемости эта группа должна встречаться реже. Так ли это в действительности?
Сифилис и группа крови 0. Перед нами стоит задача оценить возможное влияние сифилиса на дифференциальную биологическую приспособленность человеческой популяции. Как и в случае серповидноклеточности и малярии (разд. 6.2.1.6), наиболее убедительным косвенным доказательством такого влияния было бы доказательство того, что носители группы 0 имеют повышенную устойчивость к этой болезни. Это предположение в настоящее время проверить нельзя, так как сифилис настолько успешно вылечивается пенициллином, что индивидуальных различий в исходе болезни, обусловленных различием иммунного ответа, просто не существует. Однако в 1920-е годы лечение пенициллином еще не применялось. В это время был собран обширный материал по группам крови и заболеваемости сифилисом, анализ которого позволил сделать следующие выводы:
а) связь между заражением сифилисом и группами крови системы АВО отсутствует;
б) после лечения распространенным тогда препаратом неосальварсаном носители группы 0 имели гораздо большую вероятность стать «серонегативными», чем лица с другими группами крови (рис. 6.30);
в) третичный сифилис, так же как и общий паралич, реже встречается у лиц с группой 0, чем у лиц с другими группами крови системы АВО (рис. 6.31).
Итак, все данные вместе свидетельствуют о том, что группа крови 0 имеет преимущество в иммунном ответе на сифилис. Известно, что влияние сифилиса на репродукцию осуществляется в основном путем заражения плода больной матерью. Зачастую такое заражение ведет к гибели плода на поздних стадиях развития. Можно высказать предположение, что широкое распространение группы 0 у индейцев Центральной и Южной Америки обусловлено отбором за счет сифилиса и связанных с ним инфекционных заболеваний, вызываемых трепонемой.
Холера и группа крови 0 [1770]. Недавние обширные исследования холеры в Бангладеш убедительно показали существование связи между системой АВО и летально эндемичной инфекционной болезнью. Больные диареей, вызванной ротавирусом, шигеллой, токсикогенной Е. coli или
6 Популяционная генетика 333

| 
|
| Рис. 6.30.Относительная частота положительной реакции Вассермана у лиц с различными группами крови после лечения сифилиса неосальварсаном [211]. | Рис. 6.31.Относительная частота третичного сифилиса в связи с системой групп крови AB0 [211]. |
нетоксикогенными возбудителями холеры, имели частоты группы 0, сходные с контрольными (около 30%), между тем больные, зараженные токсикогенным холерным вибрионом с группой крови 0, составляли 57% Эта разница статистически высоко достоверна. Среди членов семей, зараженных токсикогенным штаммом холеры, наблюдается статистически достоверная тенденция к повышению частоты группы крови 0 с увеличением тяжести протекания диареи. Эпидемии тяжелых кишечных заболеваний, описанные в прошлом в этом регионе, вероятнее всего, были эпидемиями холеры. Низкая частота группы крови 0 в этом регионе, возможно, обусловлена более высокой восприимчивостью к холере и смертностью от нее лиц с группой крови 0. Механизм этого взаимодействия остается невыясненным
Чума и группа крови 0. Связана ли чума с распределением группы 0 в Европе? В отличие от сифилиса и холеры данные по группам крови заболевших чумой отсутствуют. Чума сейчас , болезнь очень редкая; она встречается только в областях, недоступных для исследователей. Поэтому нам придется анализировать косвенные данные.
В разд. 6.2.1.5, посвященном частотно-зависимому отбору, упоминалось, что одним из способов адаптации паразита к хозяину является выработка поверхностных антигенов, аналогичных антигенам хозяина; таким образом обеспечивается его защита от иммунного ответа. Данные, подтверждающие существование такого способа адаптации, получены для позвоночных и их паразитов [1739; 1751]. С конца 1950-х годов известно, что человек имеет общие АВН-подобные антигены со многими бактериями, особенно с бактериями группы Е. coli. Даже «нормальные» изоантитела анти-А и анти-В считаются иммунными антителами против широко распространенных кишечных инфекций. Поэтому интересно было выяснить, имеет ли Pasteurella pestis ABH-подобные антигены [1849; 1850]. Такой антиген действительно обнаружен (антиген Н). Он распространен в основном среди лиц с группой 0. Этот факт, во-первых, свидетельствует о том, что обладатели первой группы крови имеют более слабую иммунную реакцию против P. pestis и, во-вторых, согласуется с предположением о селективной невыгодности группы 0 вследствие высокой смертности ее носителей от чумы. Конечно, эти результаты нельзя считать подтверждением гипотезы.
Ослабляет ли принадлежащий микроорганизму обычный антиген группы крови иммунный ответ хозяина? Е. coli и инфекционная диарея. Ответ на этот вопрос не может быть получен на примере чумы из-за отсутствия соответствующих больных. Однако такое исследование вполне реально в отношении другой группы болезней. Они вызываются бактериями, способность которых синтезировать антигены АВН хорошо из-
334 6. Популяционная генетика
вестна; речь идет о бактериях группы Е. coli. В 1950-х-начале 1960-х годов по Центральной Европе прокатилось несколько волн инфекционной детской диареи. Возбудителем во всех случаях была Е. coli, которую путем серологического анализа антигенных спектров можно подразделить на несколько штаммов. В отличие от эпидемий прошлого исход болезни редко оказывался летальным благодаря лечению антибиотиками, переливанию плазмы.
Австрийский педиатр Киршер (1961, 1964) [1800;1801] пришел к выводу, что наиболее тяжело диарея протекает у детей, имеющих группу крови А. Этот вопрос был изучен с привлечением обширного материала, собранного в течение многих лет. Результаты оказались неоднородными: в некоторые годы большая частота заболеваемости наблюдалась у носителей группы крови А, в другие - у носителей группы 0 (рис. 6.32). Критерием тяжести протекания болезни была необходимость переливания плазмы (табл. 6.16). В те годы, когда среди пациентов преобладала группа крови А, болезнь протекала тяжелее. В годы, когда более высокую частоту имела группа 0, дети с этой группой крови болели несколько тяжелее. Эта тенденция была особенно выраженной в той части данных, где можно было идентифицировать специфический штамм Е. coli. Проведенное в это время серотипирование штаммов Е. coli показало,

| Рис. 6.32.Относительная частота групп крови А по сравнению с 0 у 1200 детей Гейдельберга, страдающих детской диареей, 1956-1963 [211]. |
| Таблица 6.16.Течение детской диареи у 396 больных с положительной реакцией на Е. coli [1889] | ||||
| Сравниваемые | 1956 и | 1960-1963 | 1957-1959 | |
| признаки | А | А | ||
| Больные, которым производилось переливание плазмы (%) | 56,6 | 39,4 | 41,9 | 52,3 |
| Потеря веса (г) | 161,0 | 137,1 | 154,0 | 158,6 |
| Средняя частота стула | 6,10 | 5,42 | 5,95 | 6,0 |
| Наивысшая температура тела (°С) | 38,40 | 38,18 | 38,51 | 38,60 |
| Время, проведенное в больнице (дни) | 26,54 | 26,13 | 23,67 | 28,10 |
| Прирост веса (г) | 577,8 | 549,0 | 506,3 | 585,6 |
что наблюдаемые различия, вероятно, связаны с соответствующей изменчивостью этих штаммов.
Ассоциация групп крови с детской диареей была продемонстрирована во многих исследованиях [1858; 1873]. Установлено, что титры антител против Е. coli 086 выше у больных с группами крови А, В и АВ по сравнению с 0 [1756] (рис. 6.33). Известно, что данный штамм Е. coli имеет антигены В и А.
Эти результаты указывают на то, что общность антигенов паразита и хозяина может действительно приводить к более тяжелой форме заболевания, если хозяином является человек, и антиген принадлежит к системе АВН. Аналогичным образом Н-антиген возбудителя чумы может привести к более тяжелому протеканию этой болезни у носителей группы крови 0. Тогда против этого аллеля будет действовать отбор.
Группы крови AB0 и оспа. Вопрос о возможной связи групп крови с оспой еще более противоречив, чем вопрос об ассоциациях, обсуждавшийся выше. Однако мы рассмотрим его здесь, поскольку данная экспериментальная схема может быть использована в качестве модели будущих исследований взаимодействия вирус — хозяин.
После разработки гипотезы о том, что распределение групп крови AB0 в популяциях человека может быть связано с крупными эпидемиями,
6. Попуяяционная генетика 335

| Рис. 6.33. Титры антител системы АВО против Е. coli 086 у лиц с различными группами крови [1756]. |

| Рис. 6.34.Относительная частота оспы у лиц с группами крови А + АВ и В + 0. Слева направо: частота (437 случаев заболевания и 428 незаболевших контрольных сибсов); 300 случаев с тяжелым и 137 случаев с легким течением заболевания; 225 больных, умерших от оспы, и 212 выживших больных; распределение групп крови среди 428 выживших больных по сравнению с 324 контрольными индивидами; тяжелые и легкие поражения кожи у 548 выживших больных [211]. |
а решающим фактором является общность антигенов возбудителя и хозяина, было решено исследовать активность АВН у возбудителя оспы. По чисто методическим причинам эти исследования провели не на вирусе натуральной оспы (вариола), а на близко родственном ему вирусе оспы коров. Была выявлена высокая активность антигена А [1849; 1890], что с очевидностью предполагает наличие иммунологического механизма, определяющего ассоциацию с данной болезнью. При попадании в организм человека вирус, имеющий антиген А, будет частично инактивирован антителами анти-А, которые присутствуют только у людей с группами крови В и 0. У лиц с группами крови А и АВ (у них этих антител нет) инактивации вируса не произойдет. Следовательно, можно ожидать, что у больных с группами крови А и АВ болезнь будет протекать тяжелее. Поскольку оспа поражает детей и часто заканчивается смертельным исходом, различия по группам крови должны оказывать сильное влияние на отбор.
Результаты, касающиеся наличия у вируса оспы А-подобного антигена вызвали сомнение [1786]. Было высказано предположение, что обнаруженный антиген А принадлежал не самому вирусу, а среде, в которой вирус выращивался. В то время когда проводилось это исследование, еще не было известно, что вирус может включать материал хозяина в собственный капсид. Сейчас получены данные в пользу существования такого механизма. Ими объясняют различную клиническую реакцию на заражение вирусом гепатита В [1818]. Вероятно, этот вирус включает компоненты белков сыворотки (особенно гамма-глобулина) и переносит их к новому хозяину, иммунная реакция которого может зависеть (по крайней мере частично) от сходства этих компонентов с его собственными белками.
Исследования ассоциации с группами крови, проведенные на больных оспой, дали противоположные результаты. Предположение, что лица, с груп-
336 6. Популяционная генетика
пами крови А и АВ будут чаще и тяжелее болеть оспой, проверялось многократно, но результаты были получены неоднозначные. В одной из работ, в которой исследовались 986 случаев оспы (давних и возникших вновь), показано, что относительная частота заболеваемости значительно выше у лиц с группами А и АВ по сравнению с В и 0 [1888]. Та же тенденция обнаружилась при анализе тяжести клинических симптомов и смертности [1888] (рис. 6.34). Более того, среди людей, переживших эпидемии оспы, наблюдался небольшой избыток обладателей групп крови В и 0, что указывает на более высокую смертность лиц с группами крови А и АВ. Среди выживших больных тяжелые поражения кожи наблюдались чаще у лиц с группами А и АВ. Это исследование было проведено в индийской деревне во время эпидемии оспы; в качестве контроля использовались сибсы пораженных пробандов, которые остались здоровыми, несмотря на то, что также соприкасались с инфекцией. Почти все жители деревни, как заболевшие, так и здоровые, никогда не подвергались вакцинации. Такая постановка эксперимента подчеркивает разницу в распределении групп крови у больных и здоровых индивидов и в то же время до минимума снижает возможность ошибки, связанной со стратификацией популяции.
Эксперименты, спланированные подобным образом, больше никогда не проводились. Однако обследование пациентов городских больниц Индии [1755; 1874] и изучение более легкой формы оспы в Бразилии [1804] не подтвердили ассоциацию оспы с группами крови. Это расхождение возможно, объясняется тем, что рассмотренное выше исследование оспы в деревнях Индии [1888] выполнено в основном на детях. В работах, проведенных в городских больницах Индии, возрастное распределение пациентов не приводится, однако на основе некоторых косвенных сведений можно предположить, что большинство больных были взрослыми.
Выше отмечалось (разд. 6.2.1.6), что селективное преимущество гетерозигот HbS характерно только для детей раннего возраста; выжившие взрослые высокоиммунны независимо от того, каким типом HbS они обладают. Вполне возможно, что это справедливо и в случае оспы. До ликвидации оспы эта болезнь в некоторых областях Индии была почти эндемической. Действительно, высокий титр препятствующих гемагглютинации антител вариолы был найден у многих жителей этих областей, никогда не имевших клинических проявлений оспы и не подвергавшихся вакцинированию [1737]. Однако данные по этому вопросу остаются противоречивыми, а возможность его окончательного решения невелика из-за практически полной ликвидации оспы.
Группа крови А и оспа в мировом населении. Как уже отмечалось, до начала 1970-х годов оспа часто встречалась во многих популяциях. По данному вопросу существуют обширные статистические материалы. Если бы оспа являлась важным селективным фактором, направленным против аллеля А, между частотой этого аллеля и частотой заболеваемости оспой или смертностью от нее наблюдалась бы отрицательная корреляция. Наличие такой отрицательной корреляции показано для популяций Индии [1727] и Африки [1885] при сравнении частоты группы А в различных субпопуляциях и частоты встречаемости в них оспы.
| Число групп популяций | Число индивидов, включенных в подсчет | Коэффициент корреляции Спирмена | |
| Индия и Па- | р = – 0,634 | ||
| кистан (смертность) | р < 0,01 | ||
| Африка (за- | 195 313 | р = – 0,499 | |
| болеваемость) | р < 0,01 |
Распределение генов групп крови AB0 среди населения земного шара и отбор за счет инфекционных заболеваний. Какие особенности распределения аллелей AB0 в мировом населении можно объяснить действием рассмотренных выше механизмов отбора, а какие нельзя? Предположительные выводы таковы:
а) исключительно высокая частота группы 0 в популяциях Центральной и Южной Америки, возможно, обусловлена преимуществом этой группы в присутствии сифилиса;
б) более высокая частота группы 0 в периферических популяциях Европы может быть вызвана более низким давлением отбора, возникающего вследствие чумы и холеры;
в) сравнительно низкие частоты группы А в Центральной и Южной Азии, возможно, объясняются отбором в результате оспы. В этих же областях частота группы крови 0 также не очень высока; соответственно повышена частота аллеля В. Это преимущество группы В может быть обусловлено длительным отбором против фенотипа А в результате оспы и
6. Популяционная генетика 337
против фенотипа 0 в результате чумы и холеры.
Если против аллеля А существует сильное и длительное давление отбора, почему группа крови А все еще присутствует в популяциях? Возможно, этот аллель имеет какое-то до сих пор неизвестное селективное преимущество. Данные относительно ассоциации групп крови с другими вирусами противоречивы [211]. Если способность включать поверхностные антигены хозяина присуща многим вирусам, частотно-зависимый отбор, возникающий в результате обычных вирусных заболеваний, может быть распространенным. Если частотно-зависимый отбор действует в пользу более редкого аллеля, вполне возможно, что эта форма отбора является основным фактором поддержания полиморфизма по системе АВО в популяциях человека.
Один из аспектов распределения аллелей системы АВО среди населения Земли не получил удовлетворительного объяснения. Почему аллель В так распространен в Центральной и Южной Азии и так редко встречается в большинстве других регионов? Частичным объяснением может быть длительный отбор как против А, так и против 0. Одним из неизвестных факторов механизма отбора по системе АВН является взаимодействие организма хозяина (человека) с кишечными микроорганизмами и, возможно, пищей, содержащей АВН-подобные антигены [1843].
Результаты популяционных исследований гена HbβE в населении Таиланда (разд. 6.2.1.7) показывают, что такие клины частот в популяциях трудно поддаются интерпретации. Они могут отражать как историю популяции и миграцию генов, так и интенсивность отбора.
Методология и результаты исследований отбора по группам крови АВО имеют значение для изучения естественного отбора в популяциях человека. Хотя несовершенство исследований эффектов отбора по группам крови системы АВО очевидно, эти работы важны с методологической точки зрения. 1. Следует признать чрезмерно упрощенным представление о том, что отбор, длительно действующий в одной и той
же популяции, является постоянным. Однако для основного фактора отбора по вариантам гемоглобина - Plasmodium falciparum- предположение о постоянстве отбора в течение многих столетий или даже нескольких тысячелетий может быть справедливым, поскольку малярия оставалась эндемическим заболеванием до тех пор, пока не изменились условия обитания ее переносчика - комара. С другой стороны, многие другие инфекционные заболевания имеют эндемичную природу. В этом случае давление отбора может измениться даже за короткий промежуток времени. Иногда возникают катастрофические вспышки заболеваний, память о которых надолго сохраняется в истории (например, эпидемии чумы в средние века). В других случаях некоторые инфекционные заболевания (такие, как детская диарея) не регистрируются вообще. Разнообразие селективных факторов и их изменение во времени - явление почти повсеместное. "
2. Генетическая изменчивость благоприятна для популяции. Если в результате эпидемии погибают почти все лица, несущие какой-либо генетический вариант, индивиды, имеющие другие варианты и поэтому менее подверженные инфекции, выживают. Следующая эпидемия может уничтожить эти варианты, но способствовать распространению первого. Таким образом, возникает динамическая ситуация, при которой генные частоты флуктуируют во времени в зависимости от преобладания того или иного фактора отбора. Данные по группам крови, полученные на костном материале XV и XVI вв., подтвердили гипотезу о колебании генных частот [1798]. Принимая во внимание трудности правильного определения группы крови АВО по ископаемому костному материалу, обусловленные иммунологической реакцией бактериальных загрязнений, к этим результатам надо относиться с осторожностью. Если благоприятной является генетическая изменчивость как таковая, единственного оптимального генотипа не существует.
3. Тот факт, что нестабильный полиморфизм благоприятен для выживания вида,
338 6. Популяционная генетика
еще на означает обязательного его поддержания. Учитывая малый размер популяций у изолированных групп на ранних этапах истории человечества, можно с течением времени ожидать перехода многих популяций к полиморфизму. Для поддержания полиморфизма необходим стабилизирующий фактор. Высказано предположение, что таким фактором может быть частотно-зависимый отбор (разд. 6.2.1.5).
Генетическая восприимчивость и инфекционные болезни. В предыдущих разделах обсуждались многие примеры отбора, возникающего в результате инфекционных заболеваний. Если в дальнейшем исследование этой проблемы будет проводиться на болезнях, вызывающих наибольшее давление отбора, оно скорее всего будет успешным. Эндемические заболевания, постоянно влияющие на популяцию, являются более эффективными факторами отбора, чем эпидемические болезни, оказывающие эпизодическое влияние. Болезни, захватывающие значительную часть популяции, более эффективны как селективные факторы, нежели те болезни, действие которых ограничивается только небольшой ее частью. Заболевания, обусловливающие гибель в раннем возрасте, служат более эффективными факторами отбора по сравнению с болезнями, не приводящими к летальному исходу или поражающими людей после окончания репродуктивного периода их жизни.
Вероятно, полиморфизм по МНС поддерживается естественным отбором, возникающим в результате инфекционных заболеваний. Логично предположить, что помимо групп крови системы АВО с инфекционными заболеваниями могут ассоциировать и другие полиморфные системы. Априорно наиболее подходящим представляется главный комплекс гистосовместимости (МНС), в особенности по генам HLA. Сведений относительно ассоциации типов HLA с основными инфекционными заболеваниями в настоящее время имеется мало. Исследователи, изучающие эту систему, в основном работают в странах, где крупные эпидемии ликвидированы или утратили свое значение. Известно что МНС является основным компонентом генетической изменчивости иммунного ответа человека (разд. 3.5.5). Следовательно, в случае HLA весьма вероятна ассоциация с эпидемическими заболеваниями и сильное давление естественного отбора. Действительно, имеются данные по ассоциации HLA с проказой [1772, 1883], тифом [1884], малярией [1852], возможно также их участие в иммунном ответе к антигенам стрептококка [1773] и столбняка [1866]. Не исключено, что в будущем отбор по системе АВО будет считаться незначительным по сравнению с отбором по МНС.
Влияет ли генетическая предрасположенность к атопическим заболеваниям на резистентность к гельминтозом (к заражению гельминтами)? Один из основных факторов, реально угрожающих здоровью детей и взрослых в тропических странах - почти поголовное заражение кишечными гельминтами, в основном ленточными червями, аскаридами и анкилостомами. Анкилостомы вызывают тяжелую анемию, которая в сочетании с другими инфекционными заболеваниями может привести к ранней гибели. Характерными клиническими признаками заражения гельминтами являются повышенное содержание эозинофильных гранулоцитов и IgE в крови. Эти же признаки характерны для атопических болезней типа астмы, сенной лихорадки и атопического дерматита. Существуют убедительные доказательства того, что атопические заболевания имеют полигенную природу и в их определении участвуют ген или гены, влияющие на уровень IgE. Атопические болезни весьма распространены в современных популяциях, вполне возможно, что генотипы, обусловливающие их, прежде обладали селективным преимуществом.
Были проведены исследования с целью установить, имеют ли атопические генотипы папуасов Новой Гвинеи [1774] селективное преимущество в отношении гельминтозов. В 5000 жителей деревни произвели кожный тест на чувствительность к нескольким аллергенам. На основании полученных результатов 10% жителей были классифицированы как обладающие атопическим фенотипом. Обследование всех пациентов местной больницы показало, что 24 из них страдают астмой. Пробы кала выявленных астматиков, 50 жителей деревни, не страдающих астмой, но имеющих атопической фенотип, и 130 жителей, не имеющих атопических признаков (контроль), были проверены на наличие яиц анкилос-
6. Популяционная генетика 339

| Рис. 6.35.Распределение числа яиц анкилостомы/грамм кала в трех выборках из сильно зараженной популяции Папуа (Новая Гвинея). Отметим огромную разницу в степени зараженности между астматиками, лицами, страдающими другими атопическими заболеваниями, и контрольной группой. |
том. Результаты представлены на рис. 6.35. Среднее число яиц гельминтов оказалось наименьшим у больных астмой, повышалось у не страдающих астмой атопических индивидов и было наиболее высоким в контрольной группе. Возможные ошибки, обусловленные различным распределением атопических и неатопических фенотипов в разных деревнях, тщательно исключались. Результаты этого исследования подтверждают выдвинутое авторами предположение: сопутствующие атопии признаки, такие как повышение уровня IgE, вероятно, обеспечивают некоторую защиту от заражения гельминтами.
– Конец работы –
Эта тема принадлежит разделу:
ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2
На сайте allrefs.net читайте: "ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Отбор по системе групп крови АВО и другим полиморфным системам
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.037 сек.
Новости и инфо для студентов