Другие формулы отбора - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2 Частотно-Зависимый Отбор [103; 1739]. До Сих Пор Мы Считали, Что Селек...
Частотно-зависимый отбор [103; 1739]. До сих пор мы считали, что селективные ценности генотипов постоянны. Однако они могут быть функцией частоты генотипов
306 6. Популяционная генетика
Рис. 6.14. Изменение частот генов по группам крови системы АВО, обусловленное отбором, направленным против детей АО и детей ВО матерей 0. р - частота аллеля А, q – частота аллеля В, r-частота аллеля 0, r стремится к 1, р и q стремятся к 0 [211].
или плотности популяции. Такой тип отбора известен как частотно-зависимый. Говоря конкретнее, при отрицательной корреляции между селективной ценностью генотипа и его частотой генотип с уменьшением его частоты становится более селективно ценным. В разд. 3.5.5 в качестве примера неравновесия по сцеплению по тесно сцепленным локусам, обусловленного селективным преимуществом некоторых комбинаций аллелей (гаплотипов), приведена бейтсовская мимикрия у бабочки Papilio memnon. Пример Papilio memnon иллюстрирует также частотно-зависимый отбор: селективная ценность мимикрирующей формы зависит от отношения ее частоты к частоте вида, который она имитирует (модели). Когда мимикрирующая форма имеет низкую частоту, хищники встречают несъедобную модель чаще, чем мимикрирующий вид. Поэтому они начинают избегать и модель, и мимикрирующую форму, что приводит к селективному преимуществу последней. Однако при увеличении частоты мимикрирующей формы хищники будут встречать ее чаще, чем несъедобную модель, и перестанут ее избегать. Если частота мимикрирующей формы намного превысит частоту модели, хищники станут связывать ее внешний вид со съедобностью. Тогда генотип станет селективно невыгодным.
Примем следующую простую модель частотно-зависимого отбора по доминантному гену:
Тогда
Следовательно, условие равновесия
w1 = w2или q2 (s1 + s2) — s1 = 0.
Данное уравнение имеет решение относительно частоты гена q тогда и только тогда, когда коэффициенты отбора s1и s2 оба положительны или оба отрицательны. Это решение имеет вид
Величина приспособленности для соотношения равновесия
Можно показать, что это равновесие стабильно в том случае, когда и s1, и s2
6 Популяционная генетика 307
отрицательны (но не меньше —1). Это значит, что стабильное равновесие существует, если аллель А имеет селективное преимущество при низкой частоте (как в случае бейтсовской мимикрии).
В состоянии равновесия приспособленности всех генотипов одинаковы. В отсутствие доминирования, если приспособленности всех трех фенотипов различны, расчеты становятся более трудоемкими [103]. Однако можно показать, что и в этом случае частотно-зависимый отбор приводит к стабильному полиморфизму при отсутствии преимущества гетерозигот.
Указанный механизм может поддерживать полиморфизм даже при селективной невыгодности гетерозигот. Более того, в некоторых случаях существует более одной точки устойчивого равновесия. Возможно, что у человека взаимная адаптация паразита к хозяину и наоборот происходит путем частотно-зависимого отбора. Паразит (например, бактерия или вирус) может адаптироваться к наиболее распространенному биохимическому или иммунологическому типу хозяина; при этом более редкие типы будут иметь селективное преимущество [1779]. Паразит имитирует антигены хозяина, либо приобретая способность продуцировать его антигены, либо прямо, используя материал мембраны хозяина для синтеза собственной мембраны. Первый механизм (генетический) реализуют бактерии, содержащие АВН-подобный антиген [211] (разд. 6.2.1.8); второй механизм, возможно, встречается у некоторых вирусов. В обоих случаях иммунные защитные системы хозяина «обмануты» и паразит размножается успешнее, чем в том случае, если бы он не имел общих с хозяином антигенов. Отбор является частотно-зависимым, поскольку вирус в основном адаптируется к наиболее часто встречающемуся генотипу и более редкие генотипы имеют селективное преимущество.
Один из способов защиты хозяина от такой «стратегии» паразита – создание им высокополиморфной системы с большим разнообразием сочетаний антигенов на поверхности клеток. В этом случае паразит не сможет получить селективного преимущества, адаптируясь лишь к одному специфическому сочетанию антигенов. Такая высокополиморфная система возникла в случае главного комплекса гистосовместимости (МНС), который включает локусы HLA, а также другие локусы, участвующие в иммунном ответе (разд. 3.5.5).
Частотно-зависимый отбор в сочетании с неравновесием по сцеплению [1739]. Отбор может зависеть не только от относительных частот генотипов в популяции, но и от отношения абсолютного размера популяции к размерам ареала, т.е. от плотности популяции. Показано, что сбалансированный полиморфизм поддерживается отбором, зависящим от плотности, в широком диапазоне условий. В определенных обстоятельствах в полиморфной популяции может присутствовать большее число особей, чем в мономорфной. Еще важнее тот факт, что изменения плотности популяции могут привести к генетическим пертурбациям, которые имитируют эффекты генетического дрейфа (разд. 6.4). Изменения популяционной плотности имели большое значение для эволюции человека, например в неолите, когда он осваивал некоторые сельскохозяйственные приемы. Поэтому можно предположить, что отбор, зависящий от плотности, играл большую роль в эволюции нашего вида.
Родственный отбор. В последнее время эволюционисты обсуждают другой тип отбора: родственный отбор (или отбор родичей: kin selection). Обычно животные взаимодействуют друг с другом и с окружающей средой, объединяясь в группы: семьи, трибы, стаи. В пределах такой группы часто наблюдается поведение, противоречащее предположению о борьбе каждой особи со всеми другими представителями своего вида за выживание и размножение, т. е. тому, что подразумевалось в ранних моделях отбора. Биологический смысл некоторых форм такого поведения (например, альтруистического), которое иногда даже стоит особи жизни, интуитивно очевиден, скажем, в случае матери, защищающей от хищника свое потомство. Но бывают случаи, когда биологическая основа поведения менее ясна, например лев, «наследующий» гарем, убивает потомство своего предшественника. Теоретическое объяснение такого социального поведения (как альтруистического, так и других типов) оставалось загадкой для эволюционистов со времен Дарвина и до тех пор, пока в 1964 г.
308 6. Популяционная генетика
Хамилтон [1784] не разработал теорию родственного отбора. Эта теория сложна и здесь будет рассмотрена только в общих чертах. Однако основная ее идея проста и очевидна: особь борется за сохранение и передачу своих генов. Часто «интересы» генов особи совпадают с ее собственными интересами; однако, к примеру, если особь имеет двух потомков, то, с точки зрения теории, пожертвовать за них жизнью равнозначно спасению своей жизни за их счет, поскольку каждый из них несет половину генов родительской особи. Если потомков трое или больше, альтруистическое поведение приобретает еще большую ценность, так как при выживании потомков сохраняется большее количество генов родительской особи, чем в случае ее собственного выживания. Аналогичные рассуждения применимы к сибсам или более отдаленным родственникам (дедам, бабкам, дядям, племянникам и т.д.). С другой стороны, лев владеет гаремом в течение ограниченного промежутка времени. Он «заинтересован» в том, чтобы за это время оставить как можно большее число потомков. Львицы, выкармливающие детенышей другого самца, не овулируют и не способны к зачатию. Следовательно, в интересах льва убить этих детенышей.
На теоретических концепциях Хамилтона и его последователей основывается социобиология – отрасль биологии, приобретающая все большее значение (разд. 7.2.4). Ее выводы стали доступными широкому кругу ученых благодаря дискуссии по поводу «эгоистических генов» и вызвали горячие споры между биологами, философами и социологами. Их понимание требует знания концептуальных предпосылок. Ниже приводится простое формальное обоснование альтруистического поведения [1738]:
Ген А обусловливает альтруистическое поведение его носителей. Например, самка дрозда, пролетая очень близко от кошки, отвлекает ее внимание от своих птенцов с риском быть пойманной самой. Альтруистическое поведение может снизить приспособленность особи на величину с, в то же время повышая приспособленность родственной ей особи на величину b. Ген А имеет селективное преимущество, если с/b < r, где r - «коэффициент связи» между донором и реципиентом, r измеряет вероятность того, что копия данного гена донора имеется у реципиента; величина r зависит только от относительного положения донора и реципиента в родословной. В отсутствие инбридинга значения r равны:
Сибсы: 1/2
Двоюродные сибсы: 1/8
Полусибсы: 1/4
Родители - дети 1/2
Это означает, что r равно доле общих по происхождению генов, которые имеют две особи (см. разд. 6.3.1).
Теперь выведем это правило для простой диплоидной модели со следующими характеристиками.
1. Один локус, два аллеля; А – ген «альтруизма» (частота равна р); а - ген «эгоизма» (частота равна q).
2. Популяция панмиктическая, ее численность бесконечно велика.
3. Ген А определяет взаимоотношения между двумя родственными особями, например «сибс—сибс» или «двоюродный сибс—двоюродный сибс».
4. Поскольку предложенное условие распространения гена А – r > с/b, то в модели условие распространения этого гена с/b < ?, и ? сравнивается с r.
Если приспособленности трех генотипов (АА, Аа и аа) равны w11 w12 и w22соответственно, условие распространения гена аллеля А [45]:
р(w11 – w21) + q (w21 – w22) > 0.
Приспособленности можно записать следующим образом:
w11 = 1 + s, w21 = 1 + r, w22= 1 + t,
где r, s и t обозначают дополнительные изменения приспособленности, происходящие в результате альтруистического поведения.
Теперь определим следующие величины: N - размер популяции (очень большой);
п - число «альтруистических действий» на поколение;
b - приращение приспособленности особи, которой была оказана помощь;
с - убыль приспособленности особи, оказавшей
помощь;
т - доля п действий в помощь индивидам АА;
е - доля п действий в помощь индивидам Аа;
х - доля п действий, где «помощник» имеет генотип АА.
Используя эти определения, можно записать приспособленности следующим образом:
В этих уравнениях генотипам просто придаются коэффициенты, определяющие уменьшение (с) или увеличение (b) их приспособленности за счет альтруистического действия (нормированные на число особей). Подставляя эти выра-
6. Популяционная генетика 309
жения в формулу 6.1, получаем
(6.11)
Конечно, знак < изменяется на обратный, если приспособленность увеличивается за счет реципиента.
Для того чтобы показать, что в случае полных сибсов (выражение 6.11) сводится к 1/2, как утверждалось выше, необходимо получить формулы для т, е и х. Они имеют следующий вид [1738]:
Если эти выражения подставить в формулу 6.11, оно сводится к 1/2. Сходные расчеты можно произвести для других степеней родства. Более общую и строгую трактовку читатель найдет, например, в работах Хамилтона.
Отбор по мультифакториальным признакам с непрерывным распределением. До сих пор мы рассматривали только отбор по отдельным генам. Однако многие нормальные и аномальные признаки и многие заболевания имеют в популяциях непрерывное распределение и детерминируются некоторым точно не установленным числом генов. В принципе, закономерности теории отбора применимы и к этим признакам. Однако, поскольку такие признаки на фенотипическом уровне пока еще не поддаются генетическому анализу (разд. 3.6), здесь необходимо использовать биометрические методы [63; 65; 124]. Эти методы имеют большое значение для селекции растений и животных. Приведем некоторые закономерности отбора по полигенным признакам:
1) изменение количественного признака в популяции под действием естественного отбора пропорционально величине наследуемости (обсуждение концепции наследуемости см. в разд. 3.6.1.5);
2) ответ признака на отбор зависит от интенсивности отбора (рис. 6.15, А и Б);
3) он также зависит от генетической изменчивости популяции (рис. 6.15). В отсутствие генетической изменчивости отбор неэффективен.
На рис. 6.16 показаны последствия искусственного отбора в нескольких поколениях. Среднее значение признака сдвигает-
Рис. 6.15.Отбор по непрерывно распределенному мультифакториальному признаку (А). Отбор происходит благодаря тому, что 50% популяции не размножается (значение признака меньше популяционной средней, незаштрихованный участок графика). В поколении Ft средняя сдвигается на s. Можно показать, что этот сдвиг равен 0,8 х стандартное отклонение (SD). (Б). Отбор происходит благодаря тому, что 80% популяции не размножается. В следующем поколении средняя сдвигается на 1,4 х SD. (В). Популяция с более низким уровнем генетической изменчивости. Если, как в случае (Б), не размножается 80% популяции, отношение изменения средней к стандартному отклонению то же самое, но его абсолютное значение гораздо ниже (в этом случае 1/2) [63].
Рис. 6.16.Действие отбора в нескольких поколениях на непрерывно распределенный признак с мультифакториальным наследованием. По мере сдвига средней изменчивость постепенно падает. То же самое происходит при действии отбора в одном поколении. В конечном итоге популяция станет генетически гомогенной и дальнейший отбор будет невозможен [63].
ся в сторону направления отбора, а генетическая изменчивость уменьшается от поколения к поколению до тех пор, пока отбор становится неэффективным.
Мутации, вызывающие слабые, почти неразличимые изменения мультифакториальных систем, вероятно, имеют большое эволюционное значение. Под действием отбора эти мутации приводят к медленным и постепенным сдвигам значений количественных признаков.
Один из наследственных признаков, имеющих непрерывное распределение и потому считающихся полигенными, - предрасположенность к таким «мультифакториаль-
310 6. Популяционная генетика
ным» болезням, как врожденные пороки развития, многие распространенные патологии или расстройства психики. Изменение давления отбора (например, путем успешного лечения врожденных болезней сердца или шизофрении) приведет к изменению кривой распределения предрасположенности к данному заболеванию и, следовательно, к увеличению частоты его встречаемости. Однако произвести количественную оценку таких изменений трудно. Мы недостаточно хорошо знаем, дает ли мультифакториальная генетическая модель адекватное описание реальной ситуации.
В предыдущих разделах рассмотрены наиболее важные математические модели отбора и некоторые их практические применения к ситуациям, встречающимся в популяциях человека. Однако мы пока не касались типа отбора, который, возможно, является основным в популяциях человека: отбора, возникающего в результате генетически обусловленных различий в восприимчивости к инфекционным заболеваниям.
Отбор в результате инфекционных заболеваний в популяциях прошлого. Естественный отбор особенно эффективен, когда он действует за счет дифференциальной смертности до достижения репродуктивного возраста, т. е в детском и юношеском возрасте. Первые достоверные статистические данные по детской смертности были получены в XVIII в. в Европе [1875]. На рис. 6.17 показана выживаемость детей от рождения до 20-летнего возраста в середине XVIII в. в Пруссии, типичной стране Центральной Европы. Более половины новорожденных не доживали до 20 лет. Приблизительно четверть из них погибала в течение первого года жизни. Каковы были причины ранней смертности7 Статистические данные отвечают на этот вопрос только частично, поскольку не все диагнозы того времени можно соотнести с известными сейчас болезнями. Однако не приходится сомневаться в том, что большинство детей умирало от вирусных и бактериальных инфекционных заболеваний, кишечных инфекций, оспы, туберкулеза и кори. Следовательно, можно априорно предположить, что инфекционные болезни являются селективными факторами. Эндемичные инфекции, воздействующие на каждое поколение, должны быть более селективно эффективными, чем эпидемии, возникающие только эпизодически, такие, например, как чума.
Риc. 6.17.Выживаемость детей от рождения до возраста 20 лет в Пруссии (Центральная Европа) в XVIII в. [1875]. Для сравнения пунктиром показана выживаемость для Берлина (1955).
На сайте allrefs.net читайте: "ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Другие формулы отбора
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
Фогель Ф., Мотульски А.
74 Генетика человека: В 3-х т. Т. 2: Пер. с англ.-М.: Мир, 1990. - 378 с, ил.
ISBN 5-03-000286-3
Книга двух известных генетиков из ФРГ и США является фундаментальным учебником
Развитие менделевской парадигмы
Разработка концепций, предложенных Гальтоном и Менделем, приблизила нас к пониманию механизма действия генов. Обсуждение близнецового метода выявило не только его возможности, но и ограничения, свя
Гены и ферменты
4.2.1. Гипотеза «один ген – один фермент»
Первые исследования. После того как в 1902 г. Гэррод указал на связь генетического дефекта при алкаптонурии с неспособност
Гены и ферменты у человека: современный уровень знаний
Круг рассматриваемых вопросов. В каждом случае ферментативного дефекта необходим особый подход в методологии исследований и интерпретации результатов. Ограниченный объем настоящего обзора за
Обнаружение и анализ ферментативных нарушений
Различия в подходах к исследованию человека и нейроспоры. Успехи в изучении ферментативных нарушений у бактерий и нейроспоры были достигнуты благодаря новому направлению исследований. Авторы
Типичные нарушения функций ферментов: ферменты эритроцитов
К настоящему времени подробно изучена группа наследственных заболеваний, связанных с недостаточностью ферментативных систем эритроцитов [933, 1345]. Эритроциты человека – безъядерные клетки, неспос
Мукополисахаридозы
Недостаточность ферментов лизосом. Ферменты или ферментные системы обычно локализуются в одном определенном районе клетки. Например, ферменты системы транспорта электронов и окислительного ф
Выявление гетерозигот
Выявление гетерозигот по ФКУ и гиперфенилаланинемии. Для заболеваний, связанных с нарушением метаболизма (в частности, ФКУ), выявление гетерозигот имеет не только теоретическое, но и практич
Необнаруженные дефекты ферментов
Сколько ферментов у человека и какие дефекты ферментов известны? Некоторые метаболические пути пока еще не выяснены. Поэтому никто не знает точного числа ферментов у человека. Согласно прибл
История изучения гемоглобина
Серповидноклеточная анемия – «молекулярное» заболевание. Изучение гемоглобина человека началось с открытия наследственного заболевания – серповидноклеточной анемии. В 1910 г. Херрик [1121] о
Генетика гемоглобина
Молекулы гемоглобина. Молекула человеческого гемоглобина состоит из четырех полипептидных цепей. Молекула гемоглобина обозначается общей формулой α2β2, котора
Генетика антител и системы антиген/рецептор
Образование антител и их функции. Живые организмы постоянно подвергаются атаке как извне - со стороны бактерий и вирусов, так и изнутри – со стороны клеток, которыев результате случайных соб
Фармакогенетика и экогенетика 4.5.1. Фармакогенетика
Развитие биохимической генетики человека, обнаружение наследственных дефектов ферментов, привели к возникновению новой отрасли генетики - фармакогенетики. Еще Гэррод, основатель биохимической генет
Механизм аутосомной доминантности
Аутосомно-рецессивные заболевания, как правило, обусловлены дефектами ферментов, которые возникают вследствие мутаций в соответствующих генах. Часто удается показать, что фермент имеет аномальную с
Аномальная агрегация субъединиц
Дисфибриногенемии (13480) [1112]. Для этой группы заболеваний характерно, что все симптомы проявляются у гетерозигот. Если такие гетерозиготы несут мутацию в гене, кодирующем белок с субъеди
Аномальные субъединицы нарушают функции мультимерных белков
Гемоглобинопатии. Существует целый ряд клинических форм гемоглобинопатии, которые имеют сходные причины возникновения, связанные с субъединичной структурой гемоглобина. Известно, что в соста
Мутации рецепторов
Рецепторы. На поверхности клеточной мембраны имеется множество рецепторов для гормонов, нейромедиаторов и лекарственных препаратов. Должно существовать множество различных мутаций, нарушающи
Наследственные дефекты клеточных мембран
Очевидно, некоторые доминантные нарушения можно объяснить возникновением мутаций, влияющих на мембраны клеток. В качестве примера приведем наследственный сфероцитоз (18290), распространенный тип ге
Доминантно наследуемые опухолевые заболевания
Существуют семьи с доминантно наследуемыми злокачественными и незлокачественными опухолями. Среди них такие часто встречающиеся наследственные заболевания, как нейрофиброматоз, полипоз и ретиноблас
Генетика эмбрионального развития
Результаты исследований по биохимической и молекулярной генетике многое прояснили в структуре генов, а также в генетическом контроле работы ферментов и других белков. В то же время наши знания о ге
Активность генов в раннем развитии
Эмбриональное развитие удобно подразделять на две фазы: раннюю, включающую оплодотворение и несколько первых делений зиготы вплоть до образования гаструлы, и позднюю, на которой закладывается форма
Поздние стадии эмбрионального развития; фенокопии
Формирование систем органов, конечностей, головы и мозга происходит на поздних стадиях эмбрионального развития. Существует ряд типичных наследственных аномалий этого этапа эмбриогенеза, что свидете
Регуляция активности генов у бактерий и эукариот
Отрицательная и положительная регуляция. Многие бактериальные гены активны только тогда, когда их экспрессия необходима, а все остальное время они выключены. Экспрессия таких генов контролир
Эффект дозы генов при трисомиях и картирование генов
Ранние работы по картированию генов с использованием эффекта дозы генов. Обычно при аутосомно-рецессивных аномалиях ферментов их активность у гетерозигот близка к величине, средней для фенот
Изучение хромосомных аберраций на уровне клеток
Фенотипы клеток при хромосомных аберрациях у человека. Биохимические исследования не позволяют сделать определенных выводов о механизмах регуляции эмбриогенеза. Вопрос можно поставить так: ч
Определение поля
Развитие половых признаков. Формирование половых признаков у человека – один из аспектов его развития, изученный относительно хорошо. Эта область представляет собой пример удачного сочетания
Спонтанные мутации
Самое важное свойство генов - их способность передаваться неизменными от поколения к поколению. Однако, если бы генетический материал никогда не менялся, была бы невозможна эволюция. Поскольку суще
Генетические изменения, обусловленные мутациями de novo
Различают следующие типы мутаций:
а) геномные мутации, приводящие к изменению числа хромосом. Геномные мутации часто возникают у растений. При этом может происходить умножение целых
Частота возникновения мутаций (скорость мутирования)
Методы оценки. Чтобы оценить частоту возникновения мутаций, необходимо подсчитать число тех случаев, когда какойлибо признак или наследственная болезнь не обнаруживаются у родителей и других
Нерасхождение хромосом и возраст матери
Статистические данные. Чем старше родители, тем больше вероятность рождения у них ребенка с синдромом Дауна. Этот факт известен уже много лет. На рис. 5.1 представлены частоты новорожденных
Генные мутации: анализ на фенотипическом уровне
Почти все трисомии, обнаруженные в популяциях человека, возникли в результате мутаций de novo. Больные с этими аномалиями появляются только в отдельных семьях; т. е. здесь мы имеем дело со «споради
Методы оценки частот мутаций
В последующих разделах будут описаны различные методы оценки частот мутаций на основе данных о редких наследственных болезнях. В большинстве случаев эти методы применяются в исследованиях, проводим
Результаты оценки частот мутаций
Оценки, полученные в популяционных исследованиях. Оценки частот мутаций приведены в табл. 5.8. Основной критерий, которым мы руководствовались, включая данные в эту таблицу,-достаточная дост
Частота мутаций и возраст отца
Блестящая идея Вайнберга. В одной из своих работ под названием "Zur Vererbung des Zwergwuchses" (О наследуемой карликовости - нем.) Вайнберг (1912) [1692] обсуждает генетические ос
Мутации
детерминирующий дистрофию Дюшенна, находится в Х-хромосоме, кроссинговер (в том числе неравный кроссинговер) может происходить только в женском зародышевом пути.
Косвенн
Генные мутации: анализ на молекулярном уровне
Анализ нуклеотидных последовательностей ДНК и аминокислотных последовательностей позволяет уточнить представление о механизмах возникновения мутаций. При этом особенно информативными оказались вари
Частоты кодонных мутаций
Какова вероятность мутирования данного кодона в определенном направлении, приводящего к замене одной аминокислоты на другую?
Первая попытка ответить на этот вопрос. Один из авторов
Проблема оценки общей частоты мутаций на геном и на поколение
Условия, необходимые для получения оценки. До сих пор мы рассматривали только частоты мутаций отдельных генов или кодонов. Было бы желательно экстраполировать эти величины и получить общую ч
Мутации в гемоглобиновых генах и генетический код
Аминокислотные замены в полипептидной цепи отражают соответствующие нуклеотидные замены в ДНК. Так как генетический код известен с начала 1960-х годов, изучение аминокислотных замен, особенн
Изучение генных мутаций в отдельных клетках
В свете успехов генетического анализа микроорганизмов представлялось многообещающим изучение проблем генетики человека на отдельных клетках. Развитие этого подхода описано в разд. 4.2.2.1. Принимая
Соматические мутации
Мутации могут происходить и в половых, и в соматических клетках. Эффект соматической мутации обнаруживается у потомков мутантной клетки, такая мутация делает индивида мозаиком. Мозаик - это особь с
Образование мозаиков по геномным мутациям
Мозаики по геномным мутациям встречаются довольно часто. Сообщалось, например, что в случае синдрома Дауна один мозаик приходится на 48 пациентов, имеющих стандартную трисомию. Исходя из оценки поп
Рак у человека с точки зрения генетики
Разнообразные данные о раке у человека позволяют сделать некоторые обобщения. Менделевское наследование раковых заболеваний человека встречается редко, однако ряд опухолей, особенно доброкачественн
Соматические мутации и старение
Старение и смерть. Люди - единственные живые существа, знающие о неизбежности смерти. Попытки примириться с этой мыслью нашли отражение в культах смерти, имеющих громадное значение в культур
Мутации, индуцированные облучением и химическими мутагенами
Общественный интерес к индуцированным мутациям. В предшествующих разделах говорилось о спонтанных мутациях. Определение «спонтанный» означает, что эти мутации происходят без какой-либо извес
Основные факты и проблемы, поставленные в ходе их анализа
Способность радиации индуцировать мутации. То, что радиация может индуцировать мутации, предполагалось с давних пор, но первые доказательства в пользу этого предположения были получены Мёлле
Облучение популяции человека ионизирующей радиацией
Насколько сильно облучается современная человеческая популяция ионизирующей радиацией? Это второй вопрос, на который должен ответить ученый, желающий оценить величину потенциального вреда, причиняе
Суть проблемы
История [1384; 1385]. То, что химические вещества индуцируют мутации, предполагалось еще на заре генетики. В своей первой публикации о радиационно-индуцированных мутациях, Мёллер (1927) [156
Популяционная генетика
Популяционная генетика отвечает на вопросы о том, как реализуются законы Менделя на уровне популяций, как влияют на генетическую структуру популяций такие факторы, как мутационный п
Закон Харди—Вайнберга: генные частоты
Закон Харди—Вайнберга в случае аутосомных генов [124]. Закон Харди—Вайнберга обсуждался в разд. 3.2. Пусть аллели А1 и А2 имеют частоты А1
Генетический полиморфизм
Определение и история вопроса. Полиморфным признаком называется менделевский (моногенный) признак, по которому в популяции присутствуют по крайней мере два фенотипа (и предположительно по кр
Наследственные болезни
Доминантные и. сцепленные с полом рецессивные заболевания. С точки зрения популяционной генетики доминантные и сцепленные с полом рецессивные болезни удобно разделить на две категории.
Систематические изменения генных частот: мутации и отбор
Частоты генов в популяциях остаются неизменными только в отсутствие возмущающих воздействий (см. равновесие Харди— Вайнберга, раздел 3.2). Наиболее важными факторами, изменяющими генные частоты, яв
Математические модели отбора: дарвиновская приспособленность
Область применения математических моделей в теории естественного отбора и их ограничения [124]. Обсуждая отбор, мы будем довольно широко использовать математические модели. В этих моделях де
Отбор, приводящий к изменению генных частот в одном направлении
Используемые символы. Приспособленность генотипа определяется как его способность производить потомство. Она измеряется не в абсолютных, а в относительных единицах, причем приспособленность
Отбор, приводящий к генетическому равновесию
До сих пор мы рассматривали только случаи отбора, приводящего к увеличению частоты одного аллеля за счет уменьшения частоты другого. Рановесие и, следовательно, стабильность генных частот в поколен
Отбор, приводящий к нестабильному равновесию
Отбор против гетерозигот. В популяции может установиться стабильное равновесие, если отбор идет в пользу гетерозигот и против гомозигот. Однако отбор может действовать и в пользу гомозигот п
История некоторых инфекционных заболеваний.
Какие инфекционные болезни могли влиять в
прошлом на частоту генов в популяциях?
1. Острые инфекционные заболевания, распространявшиеся на целые страны и уносившие значительную ча
Отклонение от случайного скрещивания
Впредыдущем обсуждении предполагалось случайное скрещивание и сохранение соотношений Харди—Вайнберга. Однако эти предположения являются абстракцией. В современных аутбредных популя
Кровнородственные браки
6.3.1.1. Коэффициент инбридинга [103]
Все люди – родственники. Родственники определяются как лица, часть генов которых общая по происхождению. Если понимать это опр
Инбридинг, размер изолята и наследственные заболевания
Частота детей с рецессивными и полигенными заболеваниями в кровнородственных и неродственных браках. Пусть аллель, который в гомозиготном состоянии приводит к возникновению рецессивного забо
Практическое применение теории
Предпринималось много попыток оценить реальный генетический груз в популяциях человека. Ранние работы представляли собой практическое приложение теории генетического груза. Однако более поздние исс
Критическая оценка
Теоретическая интерпретация. Все эти результаты, по предложению Мортона [1827], можно интерпретировать в терминах летальных эквивалентов, число которых надо определить. Однако тот факт, что
Более прямые подходы к оценке числа рецессивных генов на индивид
В последнее время используются более прямые подходы к оценке среднего числа рецессивных генов на индивид. Эти исследования ограничиваются генами, приводящими в гомозиготном состоянии к появлению ан
Дифференциация субпопуляций: генетическое расстояние
Реальная структура скрещивания популяций человека. Обычное предположение о том, что в популяциях человека преобладает случайное скрещивание - это абстракция. Выбор брачного партнера вовсе не
Поток генов
Кроме отбора (рассмотренного выше) и случайных флуктуации генных частот (которые будут обсуждаться позднее), на генофонд популяции оказывает большое влияние также поток генов. Для обозначения перен
Генетический дрейф
Детерминистические и стохастические модели. До сих пор наше обсуждение полностью основывалось на менделевских соотношениях и на законе Харди—Вайнберга. Такие популяционно-генетические параме



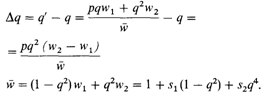



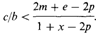 (6.11)
(6.11)









Новости и инфо для студентов