Гены и ферменты - раздел Медицина, ГЕНЕТИКА ЧЕЛОВЕКА ТОМ 2 4.2.1. Гипотеза «Один Ген – Один Фермент»
Первые...
4.2.1. Гипотеза «один ген – один фермент»
Первые исследования. После того как в 1902 г. Гэррод указал на связь генетического дефекта при алкаптонурии с неспособностью организма расщеплять гомогентизиновую кислоту, важно было выяснить специфический механизм, лежащий в основе этого нарушения. Поскольку тогда уже было известно, что метаболические реакции катализируются ферментами, можно было предположить, что именно нарушение какого-то фермента приводит к алкаптонурии. Такая гипотеза обсуждалась Дришем (в 1896 г.). Ее высказывали также Холдейн (1920 г., см. [1117]) и Гэррод (1923 г. [1091]). Важными этапами в развитии биохимической генетики стали работы Кюхна и Бутенандта [1178; 1027] по изучению окраски глаз у мельничной огневки Ephestia kuhniella и аналогичные исследования Бидла и Эфрусси на Drosophila (1936) [987]. В этих пионерских работах для выяснения механизмов действия генов были выбраны мутанты насекомых, изученные ранее генетическими методами. Однако такой подход не привел к успеху. Проблема оказалась слишком сложной, и чтобы решить ее, необходимо было:
1) подобрать простой модельный организм, удобный для экспериментального изучения;
2) искать генетическую основу биохимических признаков, а не биохимическую основу генетически детерминированных признаков. Оба условия были выполнены в работе Бидла и Татума в 1941 году [988] (см. также Бидл, 1945 [986]).
Модель Бидла и Татума. Статья этих исследователей начиналась так:
«С точки зрения физиологической генетики - развитие и функционирование организма может быть сведено к сложной системе химических реакций, которые каким-то образом контролируются генами. Вполне логично предположить, что эти гены... либо сами выступают в роли ферментов, либо определяют их специфичность. Известно, что генетики-физиологи обычно пытаются исследовать физиологические и биохимические основы уже известных наследственных признаков. Этот подход позволил установить, что многие биохимические реакции контролируются специфическими генами. Такие исследования показали, что ферменты и гены обладают специфичностью одного порядка. Однако возможности этого подхода ограниченны. Наиболее серьезное ограничение заключается в том, что при этом в поле зрения исследователей попадают наследственные признаки, не имеющие летального эффекта и, следовательно, связанные с реакциями, которые не очень существенны для жизнедеятельности организма. Второе затруднение ... заключается в том, что традиционный подход к проблеме подразумевает использование внешне проявляющихся признаков. Многие из них представляют собой морфологические вариации, основанные на системах биохимических реакций, настолько сложных, что их анализ необычайно затруднен.
Подобные соображения привели нас к следующему выводу. Изучение общей проблемы генетического контроля биохимических реакций, определяющих развитие и метаболизм, должно проводиться с помощью процедуры, противоположной общепринятой: вместо того чтобы пытаться выяснить химические основы известных наследственных признаков, необходимо установить, обеспечивают ли гены контроль известных биохимических реакций и как они это делают. Нейроспора, относящаяся к аскомицетам, обладает свойствами, позволяющими реализовать такой подход и одновременно служит удобным объектом для генетических исследований. Вот почему наша программа была построена на использовании именно этого организма. Мы исходили из того, что облучение рентгеном вызывает мутации в генах, контролирующих определенные химические реакции. Пусть для выживания в данной среде организм должен осуществлять какую-то химическую реакцию, тогда мутант, лишенный такой способности, в этих условиях окажется нежизнеспособным. Однако его можно поддерживать и изучать, если выращивать в среде, к которой добавлен жизненно необходимый продукт генетически блокированной реакции».
4 Действие генов 9
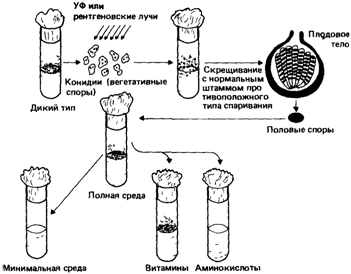
|
| Рис. 4.1. Схема эксперимента по обнаружению биохимических мутантов нейроспоры На полноценной среде мутации, индуцированные рентгеновскими лучами или ультрафиолетом, не нарушают роста гриба. Однако на минимальной среде мутант не растет. При добавлении к минимальной среде витаминов способность к росту восстанавливается При внесении аминокислот роста нет На основании этих данных можно предположить, что мутация произошла в гене, который контролирует метаболизм витамина Следующий шаг заключается в идентификации витамина, способного восстановить нормальную функцию Генетический блок обнаружен среди реакций биосинтеза витамина [1303].
|
Далее Бидл и Татум приводят описание схемы эксперимента (рис. 4.1). В состав полной среды входил агар, неорганические соли, солодовый экстракт, дрожжевой экстракт и глюкоза. Минимальная среда содержала только агар, соли, биотин и источник углерода. Наиболее подробно были исследованы мутанты, которые росли на полной среде и не росли на минимальной. Чтобы установить соединение, синтез которого нарушен у каждого из мутантов, в минимальный агар вносили отдельные компоненты полной среды.
Таким способом были выделены штаммы, неспособные синтезировать определенные факторы роста: пиридоксин, тиамин и парааминобензойную кислоту. Было показано, что эти дефекты обусловлены мутациями в специфических локусах. Работа положила начало многочисленным исследованиям на нейроспоре, бактериях и дрожжах, в которых было установлено соответствие «генетических блоков», ответственных за отдельные метаболические этапы, и специфических нарушений ферментов. Этот подход очень быстро превратился в инструмент, позволяющий исследователям раскрывать метаболические пути.
Гипотеза «один ген - один фермент» получила прочное экспериментальное подтверждение. Как показали работы последующих десятилетий, она оказалась удивительно плодотворной. Анализ дефектных ферментов и их нормальных вариантов позволил вскоре выявить такой класс генетических нарушений, которые приводили к изменению функции фермента, хотя сам белок по-прежнему обнаруживался и сохранял иммунологические свойства. В других случаях менялся температурный оптимум активности фермента. Некоторые варианты можно было объяснить мутацией, влияющей на общий регуляторный механизм и изменяющей в результате активность целой группы ферментов. Подобные исследования привели к созданию концепции регуляции активности генов у бактерий, которая включала и концепцию оперона.
10 4. Действие генов
Первые примеры ферментативных нарушений у человека. Первым наследственным заболеванием человека, для которого удалось показать ферментативное нарушение, была метгемоглобинемия с рецессивным типом наследования (Гибсон и Харрисон, 1947 [1100]; Гибсон, 1948 [1099]) (25080). В этом случае поврежденным ферментом является NADH - зависимая метгемоглобин-редуктаза. Первая попытка систематического изучения группы заболеваний человека, связанных с дефектами метаболизма, была предпринята в 1951 году. При исследовании болезни накопления гликогена [1044] супруги Кори показали, что в восьми из десяти случаев патологического состояния, которое диагностировалось как болезнь Гирке (23220), структура гликогена печени представляла собой нормальный вариант, а в двух случаях была явно нарушена. Было также очевидно, что гликоген печени, накапливаясь в избытке, не может быть непосредственно превращен в сахар, поскольку у больных проявляется тенденция к гипогликемии. Для расщепления гликогена с образованием глюкозы в печени необходимы многие ферменты. Два из них-амило-1,6-глюкозидаза и глюкозо6-фосфатаза-были выбраны для изучения как возможные дефектные элементы ферментной системы. В гомогенатах печени при различных значениях рН было измерено освобождение фосфата из глюкозо-6фосфата. Результаты представлены на рис. 4.2. В нормальной печени обнаруживалась высокая активность с оптимумом при рН 6-7. Сильное нарушение функции печени при циррозе коррелировало с незначительным уменьшением активности. С другой стороны, в случае болезни Гирке с летальным исходом, активность фермента обнаружить вообще не удалось; такой же результат был получен при обследовании второго подобного больного. У двух пациентов с менее выраженными симптомами наблюдалось значительное уменьшение активности.
Было сделано заключение, что в указанных случаях болезни Гирке с летальным исходом имел место дефект глюкозо-6-фосфатазы. Однако в большинстве более легких случаев активность этого фермента оказалась не ниже, чем при циррозе печени, и только у двух больных она была несколько меньшей (рис. 4.2).
По мнению супругов Кори, аномальное накопление гликогена в мышечной ткани нельзя связывать с недостатком глюкозо-6-фосфатазы, поскольку в мышцах этот фермент отсутствует и в норме. В качестве возможного объяснения гликогеноза мышц они предположили нарушение активности амило-1,6-глюкозидазы. Это предсказание вскоре подтвердилось: Форбс [1081] обнаружил такой дефект при одном из клинически выраженных случаев болезни накопления гликогена с вовлечением сердечной и скелетных мышц. Сейчас нам
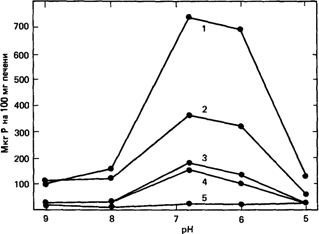
| Рис. 4.2. Освобождение фосфата из глюкозо-6-фосфата в гомогенатах печени различных больных служит мерой активности глюкозо-6-фосфатазы. 1. Значительное освобождение фосфата у больного с нормальной функцией печени. 2. Умеренное снижение при циррозе печени. 3, 4. Значительное снижение у двух пациентов с легкой формой болезни накопления гликогена. 5. Полное отсутствие ферментативной активности у пациента с тяжелой формой болезни Гирке [1044].
|
4. Действие генов 11
известно большое число ферментативных дефектов при болезни накопления гликогена [1133, 1244].
Хотя по степени проявления различные формы этого заболевания несколько различаются, в клиническом отношении между ними много общего. За одним исключением, все они наследуются по аутосомнорецессивному типу. Если бы ферментативные дефекты не были раскрыты, патология накопления гликогена рассматривалась бы как одно заболевание с характерными внутрисемейными корреляциями по тяжести течения, деталям симптоматики и срокам летального исхода. Таким образом, перед нами пример, когда генетическая гетерогенность, которую можно было лишь предполагать на основании изучения фенотипа (разд. 3.3.5), подтвердилась при анализе на биохимическом уровне: исследование ферментативной активности позволило идентифицировать специфические гены.
В последующие годы темп исследований в области ферментативных дефектов нарастал, и для 588 идентифицированных рецессивных аутосомных генов, которые Мак-Кьюсик описывает в шестом издании своей книги «Менделевское наследование у человека» (1983) [133], более чем в 170 случаях обнаружены специфические ферментативные нарушения. Наши успехи в этой области непосредственно связаны с развитием концепций и методов молекулярной генетики.
Некоторые этапы изучения ферментативных нарушений у человека. Мы приводим лишь наиболее важные вехи этого продолжающегося процесса: 1934 Фёллинг открыл фенилкетонурию
[1080]
1941 Бидл и Татум сформулировали гипотезу «один ген - один фермент» [988] 1948 Гибсон описал первый случай ферментативного нарушения при заболевании у человека (рецессивная метгемоглобинемия) [1099]
1952 Супруги Кори обнаружили недостаточность глюкозо-6-фосфатазы при болезни Гирке [1044]
1953 Джервис продемонстрировал отсутствие фенилаланингидроксилазы при фенилкетонурии [1144]. Бикель сообщил о первой попытке смягчить ферментативное нарушение, применив диету с низким содержанием фенилаланина [1004]
1955 Смитис разработал методику электрофореза в крахмальном геле [1307, 1308]
1956 Карсон и др. обнаружили дефект глюкозо-6-фосфат— дегидрогеназы (G6PD) в случае индуцированной гемолитической анемии [1030]
1957 Калькар и др. описали ферментативную недостаточность при галактоземии, показав, что у человека и бактерий наблюдается идентичное нарушение ферментативной активности [1150]
1961 Крут и Вайнберг продемонстрировали дефект фермента при галактоземии in vitro в культуре фибробластов [1177]
1967 Сигмиллер и др. обнаружили дефект гипоксантин-гуанин—фосфорибозилтрансферазы (HPRT) при синдроме Леша —Найхана [1295]
1968 Кливер описал нарушение эксцизионной репарации при пигментной ксеродерме [1035]
1970 Нейфельд выявил ферментативные дефекты при мукополисахаридозах, что позволило идентифицировать пути расщепления мукополисахаридов [1240]
1974 Браун и Голдстейн доказали, что генетически детерминированная суперпродукция гидроксиметилглютарилСоА-редуктазы при семейной гиперхолестеринемии обусловлена дефектом локализованного в мембране рецептора липопротеинов низкой плотности, который модулирует активность этого фермента (HMG) [1023]
1977 Слай и др. продемонстрировали, что маннозо-6-фосфат (как компонент лизосомальных ферментов) узнается рецепторами фибробластов. Генетический дефект процессинга препятствует связыванию лизосомных ферментов, в результате нарушается их выход в цитоплазму и последующая секреция в плазму (I-клеточная болезнь)
12 4. Действие генов
1980 При псевдогипопаратиреозе обнаружен дефект белка, обеспечивающего сопряжение рецептора и циклазы.
Все темы данного раздела:
Фогель Ф., Мотульски А.
74 Генетика человека: В 3-х т. Т. 2: Пер. с англ.-М.: Мир, 1990. - 378 с, ил.
ISBN 5-03-000286-3
Книга двух известных генетиков из ФРГ и США является фундаментальным учебником
Развитие менделевской парадигмы
Разработка концепций, предложенных Гальтоном и Менделем, приблизила нас к пониманию механизма действия генов. Обсуждение близнецового метода выявило не только его возможности, но и ограничения, свя
Гены и ферменты у человека: современный уровень знаний
Круг рассматриваемых вопросов. В каждом случае ферментативного дефекта необходим особый подход в методологии исследований и интерпретации результатов. Ограниченный объем настоящего обзора за
Обнаружение и анализ ферментативных нарушений
Различия в подходах к исследованию человека и нейроспоры. Успехи в изучении ферментативных нарушений у бактерий и нейроспоры были достигнуты благодаря новому направлению исследований. Авторы
Типичные нарушения функций ферментов: ферменты эритроцитов
К настоящему времени подробно изучена группа наследственных заболеваний, связанных с недостаточностью ферментативных систем эритроцитов [933, 1345]. Эритроциты человека – безъядерные клетки, неспос
Мукополисахаридозы
Недостаточность ферментов лизосом. Ферменты или ферментные системы обычно локализуются в одном определенном районе клетки. Например, ферменты системы транспорта электронов и окислительного ф
Выявление гетерозигот
Выявление гетерозигот по ФКУ и гиперфенилаланинемии. Для заболеваний, связанных с нарушением метаболизма (в частности, ФКУ), выявление гетерозигот имеет не только теоретическое, но и практич
Необнаруженные дефекты ферментов
Сколько ферментов у человека и какие дефекты ферментов известны? Некоторые метаболические пути пока еще не выяснены. Поэтому никто не знает точного числа ферментов у человека. Согласно прибл
История изучения гемоглобина
Серповидноклеточная анемия – «молекулярное» заболевание. Изучение гемоглобина человека началось с открытия наследственного заболевания – серповидноклеточной анемии. В 1910 г. Херрик [1121] о
Генетика гемоглобина
Молекулы гемоглобина. Молекула человеческого гемоглобина состоит из четырех полипептидных цепей. Молекула гемоглобина обозначается общей формулой α2β2, котора
Генетика антител и системы антиген/рецептор
Образование антител и их функции. Живые организмы постоянно подвергаются атаке как извне - со стороны бактерий и вирусов, так и изнутри – со стороны клеток, которыев результате случайных соб
Фармакогенетика и экогенетика 4.5.1. Фармакогенетика
Развитие биохимической генетики человека, обнаружение наследственных дефектов ферментов, привели к возникновению новой отрасли генетики - фармакогенетики. Еще Гэррод, основатель биохимической генет
Механизм аутосомной доминантности
Аутосомно-рецессивные заболевания, как правило, обусловлены дефектами ферментов, которые возникают вследствие мутаций в соответствующих генах. Часто удается показать, что фермент имеет аномальную с
Аномальная агрегация субъединиц
Дисфибриногенемии (13480) [1112]. Для этой группы заболеваний характерно, что все симптомы проявляются у гетерозигот. Если такие гетерозиготы несут мутацию в гене, кодирующем белок с субъеди
Аномальные субъединицы нарушают функции мультимерных белков
Гемоглобинопатии. Существует целый ряд клинических форм гемоглобинопатии, которые имеют сходные причины возникновения, связанные с субъединичной структурой гемоглобина. Известно, что в соста
Аномальное ингибирование ферментов по типу обратной связи и структурно аномальные ферменты
Порфирия (17600) [1282; 1217] - понижение ферментативной активности. Установлено, что различные варианты доминантной порфирии (табл. 4.25) являются следствием ферментативных дефектов
Мутации рецепторов
Рецепторы. На поверхности клеточной мембраны имеется множество рецепторов для гормонов, нейромедиаторов и лекарственных препаратов. Должно существовать множество различных мутаций, нарушающи
Наследственные дефекты клеточных мембран
Очевидно, некоторые доминантные нарушения можно объяснить возникновением мутаций, влияющих на мембраны клеток. В качестве примера приведем наследственный сфероцитоз (18290), распространенный тип ге
Доминантно наследуемые опухолевые заболевания
Существуют семьи с доминантно наследуемыми злокачественными и незлокачественными опухолями. Среди них такие часто встречающиеся наследственные заболевания, как нейрофиброматоз, полипоз и ретиноблас
Генетика эмбрионального развития
Результаты исследований по биохимической и молекулярной генетике многое прояснили в структуре генов, а также в генетическом контроле работы ферментов и других белков. В то же время наши знания о ге
Активность генов в раннем развитии
Эмбриональное развитие удобно подразделять на две фазы: раннюю, включающую оплодотворение и несколько первых делений зиготы вплоть до образования гаструлы, и позднюю, на которой закладывается форма
Поздние стадии эмбрионального развития; фенокопии
Формирование систем органов, конечностей, головы и мозга происходит на поздних стадиях эмбрионального развития. Существует ряд типичных наследственных аномалий этого этапа эмбриогенеза, что свидете
Регуляция активности генов у бактерий и эукариот
Отрицательная и положительная регуляция. Многие бактериальные гены активны только тогда, когда их экспрессия необходима, а все остальное время они выключены. Экспрессия таких генов контролир
Эффект дозы генов при трисомиях и картирование генов
Ранние работы по картированию генов с использованием эффекта дозы генов. Обычно при аутосомно-рецессивных аномалиях ферментов их активность у гетерозигот близка к величине, средней для фенот
Другие биохимические аномалии при хромосомных аберрациях
Фетальный и эмбриональный гемоглобины при трисомии 13. Нормальный синтез гемоглобина описан в разд. 4.3.2. ξ, ε и γ-цепи вместе с α-цепями входят в состав ранних э
Изучение хромосомных аберраций на уровне клеток
Фенотипы клеток при хромосомных аберрациях у человека. Биохимические исследования не позволяют сделать определенных выводов о механизмах регуляции эмбриогенеза. Вопрос можно поставить так: ч
Определение поля
Развитие половых признаков. Формирование половых признаков у человека – один из аспектов его развития, изученный относительно хорошо. Эта область представляет собой пример удачного сочетания
Спонтанные мутации
Самое важное свойство генов - их способность передаваться неизменными от поколения к поколению. Однако, если бы генетический материал никогда не менялся, была бы невозможна эволюция. Поскольку суще
Генетические изменения, обусловленные мутациями de novo
Различают следующие типы мутаций:
а) геномные мутации, приводящие к изменению числа хромосом. Геномные мутации часто возникают у растений. При этом может происходить умножение целых
Частота возникновения мутаций (скорость мутирования)
Методы оценки. Чтобы оценить частоту возникновения мутаций, необходимо подсчитать число тех случаев, когда какойлибо признак или наследственная болезнь не обнаруживаются у родителей и других
Нерасхождение хромосом и возраст матери
Статистические данные. Чем старше родители, тем больше вероятность рождения у них ребенка с синдромом Дауна. Этот факт известен уже много лет. На рис. 5.1 представлены частоты новорожденных
У какого пола и в каком из мейотических делений происходит нерасхождение хромосом?
Как было показано в разд. 2.2.1, причина трисомий - нерасхождение хромосом в мейозе. Возникают два вопроса:
5 Мутации 149
Рис. 5.4. Эффект возраста матери в сл
Генные мутации: анализ на фенотипическом уровне
Почти все трисомии, обнаруженные в популяциях человека, возникли в результате мутаций de novo. Больные с этими аномалиями появляются только в отдельных семьях; т. е. здесь мы имеем дело со «споради
Методы оценки частот мутаций
В последующих разделах будут описаны различные методы оценки частот мутаций на основе данных о редких наследственных болезнях. В большинстве случаев эти методы применяются в исследованиях, проводим
Результаты оценки частот мутаций
Оценки, полученные в популяционных исследованиях. Оценки частот мутаций приведены в табл. 5.8. Основной критерий, которым мы руководствовались, включая данные в эту таблицу,-достаточная дост
Частота мутаций и возраст отца
Блестящая идея Вайнберга. В одной из своих работ под названием "Zur Vererbung des Zwergwuchses" (О наследуемой карликовости - нем.) Вайнберг (1912) [1692] обсуждает генетические ос
Возможные различия частот возникновения мутаций у индивидов разного пола
Если большинство мутаций зависит от клеточных делений и репликации ДНК, то следует ожидать не только роста частоты мутаций в мужских половых клетках с увеличением возраста мужчин, но и абсолютно бо
Мутации
детерминирующий дистрофию Дюшенна, находится в Х-хромосоме, кроссинговер (в том числе неравный кроссинговер) может происходить только в женском зародышевом пути.
Косвенн
Герминативноклеточные и соматоклеточные мозаики по доминантным и Х-сцепленным мутациям
Анализ родословных. Если мутация происходит во время раннего развития половых клеток, возможно образование герминативных мозаиков, у которых более или менее значительная часть клеток одной и
Генные мутации: анализ на молекулярном уровне
Анализ нуклеотидных последовательностей ДНК и аминокислотных последовательностей позволяет уточнить представление о механизмах возникновения мутаций. При этом особенно информативными оказались вари
Частоты кодонных мутаций
Какова вероятность мутирования данного кодона в определенном направлении, приводящего к замене одной аминокислоты на другую?
Первая попытка ответить на этот вопрос. Один из авторов
Проблема оценки общей частоты мутаций на геном и на поколение
Условия, необходимые для получения оценки. До сих пор мы рассматривали только частоты мутаций отдельных генов или кодонов. Было бы желательно экстраполировать эти величины и получить общую ч
Мутации в гемоглобиновых генах и генетический код
Аминокислотные замены в полипептидной цепи отражают соответствующие нуклеотидные замены в ДНК. Так как генетический код известен с начала 1960-х годов, изучение аминокислотных замен, особенн
Мутации у микроорганизмов: их вклад в понимание механизма мутаций у человека
Мутации как ошибки репликации ДНК. Данные, полученные на человеке, свидетельствуют о существовании тесной связи между мутациями и клеточными делениями. Импульсом к изучению этой проблемы пос
Изучение генных мутаций в отдельных клетках
В свете успехов генетического анализа микроорганизмов представлялось многообещающим изучение проблем генетики человека на отдельных клетках. Развитие этого подхода описано в разд. 4.2.2.1. Принимая
Соматические мутации
Мутации могут происходить и в половых, и в соматических клетках. Эффект соматической мутации обнаруживается у потомков мутантной клетки, такая мутация делает индивида мозаиком. Мозаик - это особь с
Образование мозаиков по геномным мутациям
Мозаики по геномным мутациям встречаются довольно часто. Сообщалось, например, что в случае синдрома Дауна один мозаик приходится на 48 пациентов, имеющих стандартную трисомию. Исходя из оценки поп
Молекулярные механизмы хромосомной нестабильности и образование опухоли, обусловленное соматической мутацией
Пигментная ксеродерма (27870-27875). Хромосомная нестабильность и существование маркерных хромосом при трех синдромах, с нею сопряженных, наводят на мысль, что повторный разрыв хромосом може
Рак у человека с точки зрения генетики
Разнообразные данные о раке у человека позволяют сделать некоторые обобщения. Менделевское наследование раковых заболеваний человека встречается редко, однако ряд опухолей, особенно доброкачественн
Соматические мутации и старение
Старение и смерть. Люди - единственные живые существа, знающие о неизбежности смерти. Попытки примириться с этой мыслью нашли отражение в культах смерти, имеющих громадное значение в культур
Мутации, индуцированные облучением и химическими мутагенами
Общественный интерес к индуцированным мутациям. В предшествующих разделах говорилось о спонтанных мутациях. Определение «спонтанный» означает, что эти мутации происходят без какой-либо извес
Основные факты и проблемы, поставленные в ходе их анализа
Способность радиации индуцировать мутации. То, что радиация может индуцировать мутации, предполагалось с давних пор, но первые доказательства в пользу этого предположения были получены Мёлле
Проблема оценки генетического риска, обусловленного радиацией и другими мутагенными факторами окружающей среды
Проблема оценки степени риска для человеческой популяции, обусловленного радиацией и всеми другими мутагенными факторами, включает следующие вопросы:
1) Каким образом данный фактор действу
Облучение популяции человека ионизирующей радиацией
Насколько сильно облучается современная человеческая популяция ионизирующей радиацией? Это второй вопрос, на который должен ответить ученый, желающий оценить величину потенциального вреда, причиняе
Насколько может увеличиться частота возникновения спонтанных мутаций9
Как рассчитать возможность увеличения частоты спонтанных мутаций? Это третий вопрос, на который надо ответить, если мы хотим получить оценку возможного генетического повреждения, вызванного радиаци
Данные о соматических хромосомных мутациях, возникающих под воздействием радиации.
Медицинская терапия. Таф и др. (1960) [1659] первыми описали структурные аберрации в хромосомах двух пациентов, облученных при лечении анкилозирующего спондилита. С тех пор выполнено немало
Суть проблемы
История [1384; 1385]. То, что химические вещества индуцируют мутации, предполагалось еще на заре генетики. В своей первой публикации о радиационно-индуцированных мутациях, Мёллер (1927) [156
Исследовательские стратегии при оценке генетического риска, обусловленного химическими мутагенами
На какие вопросы мы должны попытаться получить ответ? Ранее мы упомянули, что перед нами поставлены четыре вопроса: как данный фактор действует на генетический материал; насколько сильно воз
Каким образом химические мутагены действуют на генетический материал?
Планирование тестирующих программ. Из предшествующего обсуждения должно быть ясно, что на первый вопрос - действует ли и как действует определенный фактор на генетический материал - мы не мо
Насколько широким является воздействие агента на человеческую популяцию?
Важный, но часто игнорируемый вопрос. Вопрос о том, насколько широко человеческие популяции подвергаются воздействию данного агента - решающий при получении любой оценки генетической опаснос
Какого увеличения частоты спонтанных мутаций, обусловленного химическими мутагенами, следует ожидать?
Химически индуцированные мутации в сравнении с радиационно-индуцированными мутациями. В разд. 5.2.1.5 мы не могли дать четкого ответа на вопрос об увеличении частоты мутаций под воздействием
Популяционная генетика
Популяционная генетика отвечает на вопросы о том, как реализуются законы Менделя на уровне популяций, как влияют на генетическую структуру популяций такие факторы, как мутационный п
Закон Харди—Вайнберга: генные частоты
Закон Харди—Вайнберга в случае аутосомных генов [124]. Закон Харди—Вайнберга обсуждался в разд. 3.2. Пусть аллели А1 и А2 имеют частоты А1
Генетический полиморфизм
Определение и история вопроса. Полиморфным признаком называется менделевский (моногенный) признак, по которому в популяции присутствуют по крайней мере два фенотипа (и предположительно по кр
Наследственные болезни
Доминантные и. сцепленные с полом рецессивные заболевания. С точки зрения популяционной генетики доминантные и сцепленные с полом рецессивные болезни удобно разделить на две категории.
Систематические изменения генных частот: мутации и отбор
Частоты генов в популяциях остаются неизменными только в отсутствие возмущающих воздействий (см. равновесие Харди— Вайнберга, раздел 3.2). Наиболее важными факторами, изменяющими генные частоты, яв
Математические модели отбора: дарвиновская приспособленность
Область применения математических моделей в теории естественного отбора и их ограничения [124]. Обсуждая отбор, мы будем довольно широко использовать математические модели. В этих моделях де
Отбор, приводящий к изменению генных частот в одном направлении
Используемые символы. Приспособленность генотипа определяется как его способность производить потомство. Она измеряется не в абсолютных, а в относительных единицах, причем приспособленность
Отбор, приводящий к генетическому равновесию
До сих пор мы рассматривали только случаи отбора, приводящего к увеличению частоты одного аллеля за счет уменьшения частоты другого. Рановесие и, следовательно, стабильность генных частот в поколен
Отбор, приводящий к нестабильному равновесию
Отбор против гетерозигот. В популяции может установиться стабильное равновесие, если отбор идет в пользу гетерозигот и против гомозигот. Однако отбор может действовать и в пользу гомозигот п
Другие формулы отбора
Частотно-зависимый отбор [103; 1739]. До сих пор мы считали, что селективные ценности генотипов постоянны. Однако они могут быть функцией частоты генотипов
306 6. Популяционная
История некоторых инфекционных заболеваний.
Какие инфекционные болезни могли влиять в
прошлом на частоту генов в популяциях?
1. Острые инфекционные заболевания, распространявшиеся на целые страны и уносившие значительную ча
Отбор по системе групп крови АВО и другим полиморфным системам
Группы крови АВО и заболеваемость. Ни одна генетическая система у человека не изучена так детально, как система вариантов гемоглобина,
328 6. Популяционная генетика
&n
Отклонение от случайного скрещивания
Впредыдущем обсуждении предполагалось случайное скрещивание и сохранение соотношений Харди—Вайнберга. Однако эти предположения являются абстракцией. В современных аутбредных популя
Кровнородственные браки
6.3.1.1. Коэффициент инбридинга [103]
Все люди – родственники. Родственники определяются как лица, часть генов которых общая по происхождению. Если понимать это опр
Инбридинг, размер изолята и наследственные заболевания
Частота детей с рецессивными и полигенными заболеваниями в кровнородственных и неродственных браках. Пусть аллель, который в гомозиготном состоянии приводит к возникновению рецессивного забо
Практическое применение теории
Предпринималось много попыток оценить реальный генетический груз в популяциях человека. Ранние работы представляли собой практическое приложение теории генетического груза. Однако более поздние исс
Критическая оценка
Теоретическая интерпретация. Все эти результаты, по предложению Мортона [1827], можно интерпретировать в терминах летальных эквивалентов, число которых надо определить. Однако тот факт, что
Более прямые подходы к оценке числа рецессивных генов на индивид
В последнее время используются более прямые подходы к оценке среднего числа рецессивных генов на индивид. Эти исследования ограничиваются генами, приводящими в гомозиготном состоянии к появлению ан
Дифференциация субпопуляций: генетическое расстояние
Реальная структура скрещивания популяций человека. Обычное предположение о том, что в популяциях человека преобладает случайное скрещивание - это абстракция. Выбор брачного партнера вовсе не
Поток генов
Кроме отбора (рассмотренного выше) и случайных флуктуации генных частот (которые будут обсуждаться позднее), на генофонд популяции оказывает большое влияние также поток генов. Для обозначения перен
Генетический дрейф
Детерминистические и стохастические модели. До сих пор наше обсуждение полностью основывалось на менделевских соотношениях и на законе Харди—Вайнберга. Такие популяционно-генетические параме
Генетический дрейф в сочетании с мутационным процессом и отбором
Мутационный процесс. Представим себе большую популяцию, состоящую из множества субпопуляций малого или среднего размера. Распределение аллельных частот q в этих субпопуляциях зависит







Новости и инфо для студентов