рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Спорт
- /
- Пассивный транспорт
Реферат Курсовая Конспект
Пассивный транспорт
Пассивный транспорт - раздел Спорт, №60.явление Диффузии. Уравнение Фика. ...
№60.Явление диффузии. Уравнение Фика.
Пассивный транспорт
-Это перенос молекул и ионов из области с большей концентрацией в область меньших концентраций (т.е. против градиента концентраций )
-Не требует затрат энергии
Виды пассивного транспорта:
а) простая диффузия
-Вещество проходит путем растворения в липидах .
-Механизмом простой диффузии происходит перенос простых незаряженных молекул (O2, CO2иN2, ), жирорастворимых веществ
-Протекает медленно
б) Диффузия через липидные белковые поры (каналы)
Порой или каналом называют участок мембраны, включающий липидные или белковые молекулы и образующий в мембране проход.
-Этот канал допускает проникновение не только малых молекул, но и более крупных ионов.
-Каналы могут проявлять селективность для(избирательность) для разных видов ионов.
в) диффузия при помощи переносчиков
Например, антибиотик валиномицин при связывании с ионом калия образует растворимый в липидах комплекс и проходит через мембрану.
Переносчики получили название ионофоры.
Диффузию через каналы или диффузию при помощи переносчиков еще называют облегченной диффузией.
Уравнения Фика или уравнение диффузии.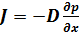
J - плотность потока вещества; D - коэффициент диффузии – м2/сек., зависит от природы вещества и численно равен количеству вещества, диффундирующего в единицу времени через единицу площади при градиенте концентарции, равном единице.
dр(знак плотности)/dx – градиент плотностизнак «-» показывает, что направление диффузии противоположно росту концентрации
Уравнение диффузии можно записать не только для плотности массового потока, но и для плотности потока частиц и плотности потока веществ, при этом в уравнении вместо градиента плотности следует использовать соответственно градиент концентрации dn/dx или градиент молярной концентрации dc/dx
_______________________________________________________________________________________
№61.Строение и модели клеточных мембран
Структура биологических мембран.
-Белки:поверхностные и интегральные (трансмембранные) -Углеводы (гликолипиды и гликопротеины) расположены на внешней поверхности… Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с…Функции мембран
· Барьерная:мембрана –селективная преграда для проникновения ионов и водорастворимых молекул
· Матричная:липидныйбислой является матрицей (структурной основой) для удержания белков и ферментов
· Механическоеразделениеклеток (или органелл) друг от друга
· Транспортная:через мембрану происходит перенос (транспорт) веществ
Модели биологических мембран
2)Бислойная липидная мембрана 3)Липосомы. Это мельчайшие пузырьки(везикулы) состоящие из билипидной мембраны… ._______________________________________________________________________________________
№64. Ионный состав цитоплазмы и межклеточной жидкости. Проницаемость клеточной мембраны для различных ионов. Разность потенциалов на мембране клетки.
Для поддержания внутреннего баланса, поступающие из внешней среды ионы должны удалятся из цитоплазмы и связываться вне клетки в нерастворимые минеральные соединения, которые не могли бы снова попасть в цитоплазму. Роль связывания ионов в цитоплазме и их транспортировки во внешнюю среду выполняют низкомолекулярные белки типа кальций-связывающих протеинов В состав цитоплазмы входят все виды органических и неорганических веществ. Например,
Натрий — участвует в поддержании мембранного потенциала, генерации нервного импульса, процессы осморегуляции и создании буферной системы крови.
Калий — участвует в поддержании мембранного потенциала , генерации нервного импульса, регуляции сокращения сердечной мышцы.
Хлор — поддерживает электронейтральность клетки
В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы — вода.
В интерстициальном внутритканевом пространстве между животными клетками находится сложное межклеточное вещество, экстрацеллюлярный матрикс. У многих тканей, например в мышцах и печени, матрикс заполняет только тонкие промежутки между клетками, тогда как в других тканях, таких, как соединительная, хрящевая и костная ткани, на межклеточный матрикс приходится большой объем и именно он выполняет основные функции . Три главных компонента межклеточного матрикса: прочные коллагены, сетчатые адгезивные белки и основное вещество, протеогликаны.
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал , а внутренняя отрицательный. В момент возбуждения полярность мембраны меняется на противоположную. В результате этого между возбужденным и невозбужденным участками мембраны возникает разность потенциалов
№65. Потенциал покоя клетки. Уравнение Гольдмана-Ходжкина-Катца
Потенциал покоя -это разность электрических зарядов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -60 милливольт.
Потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя) обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.
Основной вклад в создание и поддержание потенциала покоя вносят ионы натрия, калия и хлора.
Уравнение Гольдмана-Ходжкина-Катца:
φм=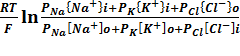
i,o- концентрации ионов вне и внутри клетки.Потенциал покоя, рассчитанный по формуле Гольдмана-Ходжкина-Катца, составляет 60мВ со знаком минус со стороны внутриклеточного пространства.
№66. Возбудимость клеток и тканей. Методы возбуждения. Закон «все или ничего».
Возбудимость клеток - способность живых клеток организма воспринимать изменения внешней среды и отвечать на них генерацией потенциала действия. Возбудимость тем выше, чем ниже пороговая сила раздражителя
Раздражители бывают: подпороговые, пороговые, надпороговые. Возбудимость и порог раздражения - обратно пропорциональные величины.
Возбуждение может быть 2-х видов:
- местное (локальный ответ);
- распространяющееся (импульсное).
Местное возбуждение возникает и в высокоорганизованных тканях под действием подпорогового раздражителя или как компонент потенциала действия. При местном возбуждении нет видимой ответной реакции.
Особенности местного возбуждения:
- нет латентного (скрытого) периода - возникает сразу же при действии раздражителя;
- нет порога раздражения;
- местное возбуждение градуально - изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя;
- нет рефрактерного периода, наоборот характерно небольшое повышение возбудимости;
- распространяется с декрементом (затуханием).
Импульсное (распространяющееся) возбуждение - присуще высокоорганизменным тканям, возникает под действием порогового и сверхпорогового раздражителей.
Особенности импульсного возбуждения:
- имеет латентный период - между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время;
- имеет порог раздражения;
- не градуально - изменение заряда клеточной мембраны не зависит от силы раздражителя;
- наличие рефрактерного периода;
- импульсное возбуждение не затухает.
Вывод:В организме животного и человека наблюдается местное и импульсное возбуждение. Возникновение того или иного вида возбуждения зависит от степени развития ткани и силы раздражителя.
Закон «всё или ничего» — правило, согласно которому на подпороговое раздражение возбудимая клетка не дает ответа, а на пороговое раздражение дает сразу максимальный ответ, причем при дальнейшем повышении силы раздражения величина ответа не изменяется.
_______________________________________________________________________________________
№67.Потенциал действия: графический вид и характеристики, механизмы возникновения и развития.
Потенциа́лде́йствия — волна возбуждении, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.
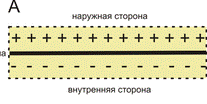

А – спокойное состояние; В –мембрана на которой возник потенциал действия
В основе любого потенциала действия лежат следующие явления:
1. Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому, что в растворе возле её внешней поверхности находится бо́льшее количество положительно заряженных частиц (катионов), а возле внутренней поверхности — бо́льшее количество отрицательно заряженных частиц (анионов).
2. Мембрана обладает избирательной проницаемостью — её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
3. Мембрана возбудимой клетки способна быстро менять своюпроницаемостъ для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю.
Третье явление является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
Фазы потенциала действия
2. Пиковый потенциал,состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны). 3. Отрицательный следовой потенциал — от критического уровня деполяризации до… 4. Положительный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной…_______________________________________________________________________________________
№68.Потенциал-зависимые ионные каналы: строение, свойства, функционирование
Для каналов характерна ионная специфичность. Каналы одного типа пропускают только ионы калия, другого — только ионы натрия и т. д.
Ионные потенциал-зависимые каналы - это каналы, которые открываются и закрываются в ответ на изменение мембранного потенциала , например, натриевые каналы , ответственные за потенциал действия Если мембранный потенциал поддерживать на уровне потенциала покоя , натриевый ток практически отсутствует, что означает, что натриевые каналы закрыты. Если теперь сдвинуть мембранный потенциал в положительную сторону и удерживать его на постоянном уровне, то потенциал-зависимые натриевые каналы откроются и ионы натрия начнут передвигаться в клетку по градиенту концентрации. Этот натриевый ток достигнет максимума и Через несколько миллисекунд ток падает почти до нуля. Закрывшись, каналы переходят в инактивированное состояние, отличающееся от первоначального закрытого состояния, при котором они были способны открыться в ответ на деполяризацию мембраны. Каналы остаются инактивированными до тех пор, пока мембранный потенциал не вернется к исходному отрицательному значению и не закончится восстановительный период длительностью в несколько миллисекунд.
При регистрации токов в очень малых участках мембраны было обнаружено, что канал открывается по принципу "все или ничего". Открытые каналы обладают одинаковой проводимостью, но открываются и закрываются независимо друг от друга, поэтому суммарный ток через мембрану всей клетки с ее многочисленными каналами определяется не степенью открытости каналов, а вероятностью быть открытым для каждого отдельного канала.
_______________________________________________________________________________________
№69.Механизм и скорость распространения потенциала действия по безмякотному нервному волокну.
Скорость проведения в нервных волокнах колеблется от 0,25 м/сек в очень тонких немиелинизированных волокнах
Распространение потенциала действия вдоль нервного волокна(аксона) обусловлено возникновением локальных токов, образующихся между возбужденным и невозбужденным участками клетки. В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал , а внутренняя отрицательный. В момент возбуждения полярность мембраны меняется на противоположную. В результате этого между возбужденным и невозбужденным участками мембраны возникает разность потенциалов, это и приводит к появлению между этими участками локальных токов. На поверхности клеток локальный ток течет от невозбужденного участка к возбужденному, внутри клетки – в обратном направлении. Локальный ток раздражает соседние невозбужденные участки и вызывает увеличение проницаемости мембран. Это приводит к возникновению потенциалов действия в соседних участках. В то же время, в ранее возбужденном участке происходят восстановительные процессы реполяризации Вновь возбужденный участок в свою очередь становиться элекроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс повторяется многократно и обусловливает распространение импульсов возбуждения по всей длинне клетки в обоих направлениях. В нервной системе импульсы проходят лишь в определенном направлении из-за наличия синапсов, обладающих односторонней проводимостью.
Удельное сопротивление биомембран велико, но вследствии их малой толщины сопротивление изоляции в сотни тысяч раз меньше, чем у технического кабеля.По этому однородное нервное волокно не может проводить электрический сигнал на далекие расстояния.
λ=корень из (dR/4р)
d- диаметр волокна, R - поверхностное сопротивление мембраны в Ом * м2 и р-удельное сопротивление аксоплазмы в Ом*м.
С увеличением λ (постоянная длины) степень затухания сигнала уменьшается, при этом возрастает скорость проведения импульса. Увеличения постоянной длинны λ можно добиться путем увеличения диаметра d аксона.
_______________________________________________________________________________________
№70.Механизм и скорость распространения потенциала действия по миелинизированному нервному волокну.
У высокоорганизованных животных затухание сигнала предотвращается с помощью миелиновой оболочки вокруг аксона. Примерно через каждые 1-3 мм вдоль миелиновой оболочки имеется перехват Ранвье.
Центральной его частью является аксон, по мембране которого проводится потенциал действия. Аксон заполнен аксоплазмой — вязкой внутриклеточной жидкостью.
С увеличением λ степень затухания сигнала уменьшается, при этом возрастает скорость проведения импульса .
Удельное сопротивление миелина значительно выше удельного сопротивления других биологических мембран.кроме того толщина миелиновой оболочки во много раз больше толщины обычной мембраны, что приводит к возрастанию диаметра волокна и соответственно величины постоянной длины . λ
В связи с большим сопротивлением миелиновой оболочки по поверзности аксона токи протекать не могут. При возбуждении одного узла возникают токи между ним и другими узлами. Ток подошедший к другому узлу, возбуждает его, вызывает появление в этом месте потенциала действия, и так процесс распространяется по всему волокну. Затраты энергии на распространение сигнала по волокну, покрытому миелином значительно меньше чем по немиелинизированному.
– Конец работы –
Используемые теги: Пассивный, транспорт0.05
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Пассивный транспорт
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.108 сек.
Новости и инфо для студентов