рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- Эволюционные теории асимметризации организмов, мозга и тела
Реферат Курсовая Конспект
Эволюционные теории асимметризации организмов, мозга и тела
Эволюционные теории асимметризации организмов, мозга и тела - раздел Философия, Удк 612.82 Эволюционные Те...
УДК 612.82
Эволюционные теории асимметризации организмов, мозга и тела.
В.А. Геодакян
E-mail: geod@newtech.ru; тел. (095) 137-65-76 д.
Институт проблем экологии и эволюции им. А.Н.Северцова, РАН, Москва
“Не на основе фактов строятся теории, как
думают представители индуктивной науки, всегда
на основе теории факты укладываются в систему”
А.А.Любищев
“Если не имеешь в голове идеи, – не увидишь фактов”
И.П.Павлов
“Представляется, что человеческий разум должен
свободно строить формы, прежде чем подтвердится их
реальное существование... Из голой эмпирии не может
расцветать познание. Такой расцвет возможен только
из сравнения придумываемого и наблюдаемого” .
А.Эйнштейн
Аннотация
Отношение степени изменения и сохранения – главная характеристика эволюции систем. Для мономодальных (унитарных) систем (бесполых, симметричных), они альтернативны. Чем больше одно, тем меньше другое. Дифференциация на две сопряженные подсистемы, специализированные по консервативным и оперативным аспектам эволюции, позволяет, улучшая одновременно обе, повышать устойчивость системы. Это достигается дихронной эволюцией, при которой оперативная подсистема эволюционирует раньше консервативной. Так, эволюция мужского пола опережает женского, а левого полушария мозга – правого. Эволюционный дихронизм создает эволюционный диморфизм. В развитие, созданной ранее, эволюционной теории функциональной асимметрии мозга и рук, предлагается новая концепция явлений правшества и левшества, как цис-транс асимметрии особей, аналогов мужского и женского пола. Ее основные положения. 1. В филогенезе симметрия организмов неуклонно падает, асимметрия растет: шаровые, радиальные, билатеральные и “венец” эволюции – триаксиально асимметричные. Последняя ось асимметризации – латеральная. 2. В отличие от существующих теорий, рассматривающих правшество – нормой, а левшество – патологией, следствием аномалий развития или родовой травмы, новая концепция трактует их, как нормальные, адаптивные фенотипы для стабильной и изменчивой среды, регулирующие поведенческую пластичностьобщества. 3. В эмбриогенезе доминантное полушарие, как генотип, реализует доминантный фенотип парных органов: в оптимальной среде – в транс позиции, в экстремальной – в цис и создает новый, позиционный вид асимметрии для организма и популяции. 4. Транс и цис фенотипы реализуют половые гормоны. Поэтому 5. У трансособей шире норма реакции, ýже фенотипическая дисперсия и выше адаптивность в стабильной среде; у цисособей, наоборот, ýже норма реакции, шире фенотипическая дисперсия и выше адаптивность в изменчивой среде. 6. Как новый ген возникая в мужском геноме, проходит там проверку в течение многих поколений, после чего попадает в женский, так и центры управления новых функций, возникая в левомполушарии, после проверки транслоцируют в правое. 7. Значит, в левом полушарии (социальном) локализованы центры эволюционно молодых функций, в правом(биологическом) – старых. 8. Доминантность полушария (парного органа) в фазах эволюции означает наличие или избыток информации по данной функции, а субординатность – отсутствие или нехватку. 9. В стабильных фазах (симметричных), при отсутствии функции доминирует левое,поисковое полушарие, при наличии функции, более совершенное – правое. 10. Как доля мужских особей определяет эволюционную пластичность популяции (генотипов), так и доля цисособей определяет поведенческую пластичность общества (фенотипов, психики). 11. Экстремальные условия смещают равновесия [♂♂]«[♀♀] и [цис]«[транс]: в эмбригенезе влево, в постнатальном онтогенезе вправо, повышая “оборачиваемость” ♂♂ и цисособей. Это осуществляют андрогены, приближающие систему к среде, или эстрогены, удаляющие от нее.
1. Введение.
Всякий раз, когда науке удавалось объяснить, казалось бы, совершенно разные явления с единой позиции, достигался существенный прогресс. Много свидетельств тому в теоретической физике. Принцип скорейшего пути Фермà в геометрической оптике свёл воедино законы распространения, отражения и преломления света. Ньютоновская механика и теория гравитации объединили теорию свободного падения тел Галилея и законы движения планет Кеплера. В свою очередь, три закона Ньютона были сведены к принципу наименьшего действия Мопертюи. Теория Максвелла связала в единое целое не только электрические и магнитные явления, но и оптические и свела их к 24 уравнениям, которые в дальнейшем Герц и Хевисайд сократили до четырех, а Эйнштейн – до одного. Такая унификация – закономерный процесс. В теоретической биологии он еще предстоит.
Казалось, что общего между такими разными явлениями, как пол и асимметрия? Разделение популяции на женский–мужской пол, генома на аутосомы–гоносомы и общества на правшей–левшей, организма, парных органов (мозга, рук) на, отличающиеся во многом друг от друга, левые–правые половины?
В эволюционном плане они функционально изоморфны: имеют общие цели (предназначение), закономерности и механизмы. Все они – бинарные сопряженные дифференциации (БСД)[1], возникшие в процессе эволюции из унитарных, мономодальных форм (нераздельнополых, симметричных). Это – адаптивно эволюционирующие, следящие системы, осуществляющие особую, экономную форму информационного контакта со средой. Все они состоят из консервативной и оперативной подсистем, возникающих в результате дихронной эволюции, регулирующих пластичность системы для эффективной эволюции в изменчивой среде. Поэтому, вскрыв эволюционную логику одной из них, по аналогии можно объяснить и другие.
Эти проблемы, как эволюционные, не решаются в плоскости неэволюционных наук, оперирующих малыми масштабами времени. Поэтому, заранее обречены на неуспех все попытки решить загадку асимметрии в рамках, эмбриологии, патологии или аномалий развития, точно так, как до сих пор безуспешны попытки решить проблему пола, как репродуктивную. В этой связи, на Западе уже четверть века пишут, о “кризисе” в эволюционной теории: “Поразительно, но ученые не могут убедительно ответить, для чего существует пол” (Crews, 1994). Это неверно! Ответ на этот вопрос был опубликован в статье в математическом журнале, после того, как ее отклонили биологические (Геодакян, 1965).
Идея асинхронной эволюции, предложенная мной для понимания эволюционной роли дифференциации полов, оказалась очень плодотворной и привела к созданию эволюционной теории пола. Впервые удалось объяснить адаптивное значение раздельнополости, ее преимущества перед другими способами размножения, вскрыть эволюционные закономерности и механизмы (Геодакян, 1965, 1989, 1991). Потом я обобщил эту идею для трактовки аналогичных структур целого ряда БСД от молекулярного уровня организации до популяционного, социо-культурного и технического (Геодакян, 1972).
На такой общей основе были созданы, помимо ЭТП, изоморфные эволюционные теории, объясняющие функциональную асимметрию мозга и видовую праворукость человека (Геодакян, 1980, 1984, 1986, 1987, 1992, 1993), обладающие исключительным для биологических теорий объяснительным и предсказательным потенциалом. Позже такая трактовка была распространена на хромосомную дифференциацию генома (Геодакян, 1996, 1998, 2000) и на явление леворукости (Геодакян, Геодакян, 1997).
Цель данной статьи – в развитие, полученных ранее выводов, на основе той же идеи и, аналогии пол–асимметрия, предложить внутренне непротиворечивую концепцию правшества–левшества, включающую высшие (поведенческие и психологические) проявления феномена на уровнях организма и популяции. Идея дихронизма способна объясниь с единых позиций, максимум известных фактов и успешно предсказывать новые. Она впервые позволяет понять адаптивность левшества и его тесную связь с полом (главные камни преткновения для всех теорий), закономерности реализации, механизмы регуляции, вскрыв, эволюционные корни явления, связать его с условиями среды, поведением, психологией и ответить, как минимум, на следующие общие вопросы. 1.Что такое латеральная асимметрия полушарий, леворукость, левша, левшество? Норма или патология? 2. Имеют ли они адаптивное значение, какое? 3. Какой принцип лежит в основе функциональной специализации полушарий мозга и других парных органов? Что они дают? 4. Когда и как появляются они в филогенезе и онтогенезе? 5. Как они связаны со средой, полом, генотипом, нормой реакции, гетерозиготностью, гормонами, поведением и психикой? 6. Что такое доминантность? 7. Каковы конститутивно-факультативные отношения между D-полушарием и D-рукой? 8. Для чего мозолистое тело? 9. Какова роль контра и ипси нервных связей полушарий мозга с левой и правой сторонами тела (парными органами)? А также многие частные вопросы, например, 1. Почему вербальные функции, локализованы в левом полушарии и лучше развиты у женщин, а пространственно-зрительные – в правом и лучше у мужчин? 2. Почему у человека видовая праворукость и левоногость, сердце смещено влево, а зубы слева крупнее, чем справа? 3. Почему афазия и апраксия у женщин чаще при поражениях передних отделов мозга, а у мужчин – задних? 4. Почему у дельфинов действует (овулирует) только левый яичник, у кашалотов – левая ноздря, у новозеландских куликов клюв загнут вправо, а у бассетов и спаниелей самое длинное ухо должно быть правое у самцов, а самое короткое – левое у самок? И т.д. и т.п.
Термины, сокращения и обозначения. Латеральная асимметрия проявляется на четырех уровнях организации: отдельных функций, парных органов, организма и популяции. Их векторы доминантности обозначены полужирными буквами, а сокращения курсивом. Поэтому, ЛП, ПР следует читать “левополушарность”, “праворукость”, а ПП, ЛР – “правое полушарие”, “левая рука”. С функциями и органами все ясно и однозначно: лево (право) полушарность, рукость, ногость и т.д. Термины левша–правша часто применяют как синонимы рукости. Считаю, неправильно: это более обобщенные, комплексные понятия, которые следует отнести к целому организму, с учетом не только рук, но и других парных органов; а главное, к высшим проявлениям латеральной асимметрии по поведенческим и психологическим признакам организа, отражающим её адаптивность в разных средах. Правшиэто особи у которых D-полушарие и D-рука находятся в транс позиции (с разных сторон тела), поведение которых более адаптивно в стабильнойсреде. Левши это особи, имеющие D-полушарие и D-руку в цис позиции (с одной стороны тела), поведение которых более адаптивно в изменчивой среде. Еще более обобщенные понятия левшества–правшества, применяемые, как названия феномена в целом или множеств, следует отнести к популяционному уровню, для характеристики, асимметрии численности, дисперсии и модальности субпопуляций левшей и правшей. Они определяют эволюционную пластичность общества на уровне поведения и психики. Это аналоги, соответственно, соотношения полов, дисперсии полов и полового диморфизма.
Проблема: факты, загадки, существующие теории.
Мозг у позвоночных – парный орган. Его разделение на два полушария обусловлено билатеральным планом строения тела, симметрией сенсорных и моторных… Считается, что все парные органы человека (легкие, почки, яичники, семенники)… Левое полушарие (у нормальных праворуких) служит для смыслового восприятия и воспроизведения речи, письма, тонкого…Теоретические основы новой концепции.
Тогда, среди живых систем должно быть много БСД. Их действительно очень много. ГЦ–АТ пары нуклеотидов, COOH–NH2 группы аминокислот, двойная спираль… Они прослеживаются и в антропогенной формации, социокультурных, технических,… БСД корнями уходят в неживую формацию: они аналоги факторов экстенсивности–интенсивности (обобщенных…Эволюционный ряд основных типов морфологической симметрии организмов.
4.1. Симметрия и эволюционный прогресс. Организмышаровой, радиальной и билатеральной симметрии широко распространены в животном мире и хорошо известны.…Если эволюция идет в сторону появления или улучшения функции, то доминантной будет её авангардная форма. Если же функция эволюционно регрессирует: утрачивается или ухудшается (напр., обоняние, хватательный рефлекс у человека), то – арьергардная форма (рис. 5).
5.5. Общая трактовка полового и латерального диморфизма, доминантности и асимметрии.
Идея дихронной эволюции вскрывает полную аналогию в эволюции пола, мозга и генов(рис. 5). Их филогенез состоит из одинаковых, чередующихся во времени стабильных и мономорфных (1,3,5), и, эволюционных и диморфных (2,4) стадий, т.е. и половой диморфизм и латеральный возникают только при эволюции, в результате дихронизма. Вторая общая (для всех БСД) особенность – разная дисперсия фенотипов (ОП>КП), как следствие разной нормы реакции (КП>ОП). Это говорит о том, что должна существовать: 1. Общая гормональная основа для определения половых и латеральных фенотипов; 2. Общая трактовка доминантности центров функций в полушариях и D генов в диплоидном геноме, как диморфизмов.
Ведь обе пары М–Ж и ЛП–ПП ведут себя в стадиях эволюции совершенно одинаково. Их “одноколейный” путь превращается в “двухколейный”, образующий “петли” дихронизма(окрашены). По изменеию площади которых со временем можно судить: кто, когда, сколько и откуда получает новой информации, кто за нее платит и кому? А по направлению пути, по углу отклонения (a) от вертикали, можно судить о скорости эволюции (v = tga) и дисперсии фенотипов, которая растет с ростом скорости и приводит к появлению диморфизмов (полового и латерального). А так как дисперсию определяет норма реакции, а последнюю половые гормоны, то можно сделать вывод, что половые гормоны определяют и латеральные фенотипы.
Согласно ЭТП, пол – экономная форма информационного контакта со средой для эффективной эволюции. Тогда половые гормоны, регулирующие интенсивность этого контакта, а так же цис–транс фенотипы, приобретают новый экологический и эволюционный смысл.
Эстрогены, как стабилизирующие гормоны, нормализующие распределение фенотипов (сужающие дисперсию), действующие центростремительно, удаляющие систему от среды и замедляющие эволюцию. А, определяемые ими транс особи, как “стабильное ядро”, эволюционный арьергард (аналог женского пола).
Андрогены, как дестабилизирующие гормоны, аномализирующие распределение фенотипов (расширяющие дисперсию),действующие центробежно, приближающие к среде и ускоряющие эволюцию. А, определяемые ими цис особи – “лабильная оболочка”, эволюционный авангард (аналог мужского пола) (Геодакян, 2000)
По теории БСД, широкая норма реакции(пластичность в онтогенезе, Геодакян, 1974), удаляя КП от среды, превращает её в более устойчивую, ценную и совершенную подсистему (Ж, ПП), а узкая норма реакции ОП, напротив, приближая к среде, превращает её в “буферную”, более “дешевую”, поисковую подсистему (М, ЛП).
По теории пола, генотипический половой диморфизм, gsd =IМ – IЖ = DI =Iп,
где: IМ, IЖ – количество генетической информации в геномах мужского и женского пола, DI – их разность, Iп – информационный потенциал gsd (Геодакян, 1972, 1975, 1998б), т.е. появившийся в дивергентной фазе под действием экологического потенциала, оплаченный мужским полом, gsd служит для женского пола фактором “бесплатной” (минуя отбор) эволюции. В этом адаптивный смысл пола (Геодакян, 1965, 1972, 1974, 1989, 1991, 2000).
По аналогии, латеральный диморфизм, ЛД = IЛП – IПП = DI = Iп = D = Ас,
где: IЛП, IПП – количества информации в левом и правом полушарии, DI – их разность, Iп – информационный потенциал ЛД, D – доминанность, Ас – асимметрия.
Это значит, что, все 4 явления (gsd, ЛД, Ас и D), не что иное, как некие потенциалы, факторы эволюции, движущие более ценные, но инерционные подсистемы (Ж пол и ПП).
Теория предсказывает конкордантность векторов ПД и ЛД по любой функции. Например, хватательный рефлекс (или обоняние, угасающие признаки) д.б. максимален у девочек слева и минимален у мальчиков справа. А ширина лопаток (чел.) или длина ушей у бассетов и спаниелей (новые признаки), наоборот: максимальны у мальчиков справа и минимальны у девочек слева. Общая формула для всех таких тетрад: (ПМ + ЛЖ)/2 » ЛМ » ПЖ, где: ПМ, ЛЖ, ЛМ, ПЖ – количественные признаки у мужского и женского пола, справа и слева.
В стадиях эволюции (2,4) доминирует всегда наличие или избыток информации (DI), при появлении или улучшении функции (2), это авангардная форма, а при утратеили ухудшении (4), – арьергардная (напр., обоняние у человека). Это главный критерий доминантности. А в стабильных (1,3,5) стадиях, при равенстве количеств информаций, доминантность определяют динамические характеристики полушарий, т.к. при симметричном отсутствии функции (1,5) – важнее её поиск,а присимметричномналичии(3) – её совершенство, то в первых случаях доминирует инновационно-поисковое левоеполушарие, а во-втором – более совершенное (быстрое) правое (рис.5). Действительно, скорости процессов в правом полушарии выше, чем в левом (Костандов, 1978), даже плотность правого полушария больше, чем левого (Walker, 1980). И то, и другое, видимо, связано с совершенством ПП.
Как видно из графика, информационный компонент для полушарий однозначно определяется логикой дихронной эволюции, полностью аналогичной эволюции доминантности генов и признаков у мужского и женского пола. Стало быть, направление вектора доминантности полушарий определяется вектором латерального диморфизма: он направлен от отсутствия или нехватки информации к её присутствию (окраска темнее) или избытку(светлее), а степень доминантности пропорциональна диморфизму (половому или латеральному) (рис. 5).
Таким образом, полный цикл эволюции, появления и утраты функции, состоит из следующих последовательных стадий: 1. Доэволюционная стабильная стадия симметрично “пустых’’ (по данной функции) полушарий (аналог рецессивной гомозиготы, аа); 2. Эволюционная стадия появленияфункции, информационная асимметрия полушарий (аналог гетерозиготы, аА); 3. Межэволюционная стабильная стадия симметрично “полных” полушарий (аналог D гомозиготы, АА). 4. Эволюционная стадия утраты функции, информационная асимметрия полушарий (аналог гетерозиготы, аА); 5. Послеэволюционная стадия симметрично “пустых” полушарий (аналог рецессивной гомозиготы, аа).
Конечно, в случае генов имеем дело с линейной информацией, тогда как в полушариях мозга, она может быть двумерная (об этом говорит развитая поверхность коры: наличие извилин и борозд), или даже трехмерная (голографическая), но приведенные соображения остаются в силе и в этих случаях.
Аналогия ЛП с мужским полом и ПП – с женским не ограничивается только стадиями эволюции (2,4), но прослеживается и для стабильных (1,3,5) когда D определяет поиск или совершенство, а также и в психологии.
В фазе поиска и освоения новых функций лучше М пол и ЛП, которые освоив их, передают Ж полу и ПП, а в фазе совершенства начинают доминировать последние. Так во всем: в скорости и беглости речи (выше у Ж!), по почерку, качеству работы на конвейере и т.д. Есть и эксперименты, подтверждающие это (Landauer, 1981).
Пребывание центров новых функций в левом полушарии (генов в М геноме, “карантинная служба”) необходимо для апробации и отбора. А при утрате старых функций (генов) их утрачивает сначала левое полушарие (М геном), а потом правое (Ж геном, “архивная служба”) на случай возврата старых условий среды, рис. 5 (Геодакян, 1993, 2000).
Изоморфизм БСД (генотип–фенотип, Ж–М, ПП–ЛП), управляющих и управляемых подсистем в генетике (родители–дети) и нейробиологии (полушария–руки) позволяют трактовать полушарность, как генотип, а рукость, как фенотип. Эта межпарная асимметрия у нас наследство от билатерально симметричныхорганизмов.
Появление внутрипарной асимметрии у триаксиально асимметричных форм, превращает симметричные полушария и руки в два вектора: полушарности ирукости. Если они дискордантны, то доминантное полушарие и доминантная рука будут в транс позиции (на разных сторонах тела), если конкордантны – в цис позиции (на одной стороне), т.е. возникает 4 градаций D-S органов. На рис. 6 доминантные органы пары изображены темнее и чуть выше, чем S органы. Цвет наглядно показывает градации D-S, а высота – конфигурацию D-S.

Реликтовая асимметрия, полушарность ирукость создают у человека 2 генотипа: D-ЛПили D-ПП, а они, в свою очередь, по 2 фенотипа, имеющих две разные друг от друга и от билатерально симметричных организмов, конфигурации D-S. У опоссума – квадрат(П=Р=0), у транс особей – трапеция(П, Рдискордантны) у цис особей – ромб(П, Рконкордантны), выявляющие уникальные свойства цис структур: сверх D (повышенное артериальное давление D полушария от D руки в цис позиции) и сверхобучение (max Iп между D полушарием и S рукой по контра связи, рис. 6).
5.6. Проверка теории. Объяснение и предсказание фактов.
Гипотеза транслокации D и, вытекающий из нее, “возрастной принцип” локализации функций по полушариям позволяют объяснить с единых позиций подавляющее большинство известных фактов и явлений и успешно предсказывать, ниоткуда иначе не следующие, новые.
Совершенно очевидно, что большинство левополушарных функцийявно эволюционно моложе, чем правополушарные(см. 2.1.). Но есть и такие, эволюционный возраст которых не так очевиден. Например, эмоции: отрицательные–положительные; понимание: пространства–времени, существительных–глаголов, пространства цветов; способность высказываний: истинных–ложных. Как в филогенезе, так и в онтогенезе первые появляются раньше вторых. У новорожденных детей плач предшествует улыбке, у котят жалобный писк опережает мурлыканье, у щенят скуление начинается на три месяца раньше виляния хвостом. Кроме того при функциональном угнетении мозга отрицательные эмоции исчезают последними и восстанавливаются первыми (Денисова, 1978), что означает более древний возраст. Если вспомнить мышление и лексику детей или представителей молодых культур (дикарей), то легко убедиться, что понимание пространства проще, чем времени; существительных проще, чем глаголов; истинные высказывания проще, чем ложные. Ориентация в пространстве приходит раньше, чем во времени; первые слова ребенка – чаще существительные, лукавство и ложь появляются позже. Пространство цветов, также, видимо, можно считать эволюционно новым приобретением. Персонажи Гомера пользовались очень узким спектром названий цветовой гаммы (конечно, если это не связано с его слепотой).
Интересна ситуация с музыкой и юмором. Считалось, что их центры в правом полушарии (Денисова, 1978). А по новой теории там д.б. их старые, “нетворческие” версии, а творческие д.б. в левом, об этом говорит избыток мужчин среди композиторов и юмористов (Геодакян, 1987). Наши предки слушали пение птиц, пытались подражать, научились петь, в этом смысле музыка – эволюционно старая функция и должна управляться правым полушарием. Но музыкальное творчество (как и любое другое) – новая функция и её центр должен находиться в ЛП (Геодакян, 1987, 1993, Harris, 1978). Так трактует новая теория известные факты. Однако гораздо убедительнее, проверить, вытекающие из нее предсказания.
1. Парные органы билатерально симметричных форм возникали, видимо из непарных. И те и другие вначале д.б. симметричны, а потом становиться асимметричными; при этом, старые функции должна сохранять за собой, как правило, консервативная подсистема, субординатный орган (чаще левый), новые приобретать – оперативная, доминантный орган (чаще правый). Этот вывод теории подтверждается. Например, у дельфинов овулирует, в основном, левый яичник; у кашалота: дышит только левая ноздря, правая закрылась, и превратилась в пазуху для сжатия воздуха, при рекордном (>2 км!) нырянии (Томилин, 1957). Кстати, это правило объясняет, почему сердце человека слева. При переходе далеких наших предков от жаберного дыхания к легочному, правое легкое, появившись первым, оттеснило, исходно по центру расположенное, сердце влево. А через много поколений (время равное дихронизму), когда возникало левое легкое, место уже частично было занято. Поэтому правое легкое из трех долей, левое – из двух. Другого объяснения я не смог найти (Геодакян, 1993,1998 б). Еще один пример, изогнутый вправо клюв, появившийся для вскрытия раковин моллюсков у новозеландских куликов.
У триаксиально асимметричных форм, в процессе эволюции, происходит дальнейшая асимметризация, которая, приводит к превращению парных органов в разные.Поэтому уместна даже гипотеза о таком происхождении селезенки (с) и печени (п): расположены на одном уровне, имеют много общих функций (jоб), наряду с частными (jс и jп). Если окажется, что jоб древнее jп, но моложе jс, или же у мужчин jп > jс, а у женщин – наоборот, то гипотеза верна!
2. Потоки анизотропной информации от среды создают в организме и полушариях мозга её градиенты, тогда триаксиальная асимметрия, накладываясь на фон реликтовой, должна распространяться спереди назад. Действительно, черная окраска спины, у изначально белых пингвинят, появляется на голове, потом разливается к хвосту. Такая же картина у китов: детёныши одних видов темнеют с головы (серые киты), других светлеют (белухи) (Томилин, 1957), при этом, и там и там, правая сторона должна опережать левую.
3. Д. Кимура сделала загадочное открытие: афазия и апраксия у женщин – чаще следствие поражения переднего отдела мозга, у мужчин – заднего (Кимура, 1992). Оно объясняется наложением дихронизмов: по полу и по оси лоб–затылок (центры, которые у женщин еще в переднем отделе, у мужчин уже в заднем).
4. Другое интересное подтверждение наложения двух дихронизмов – расположение гонад по сегментам тела у дождевых червей. Поскольку у билатерально симметричных организмов информация идет от головы к хвосту, а яичники древнее семенников, теория предсказывает, что они должны быть ближе к хвосту, чем семенники. Оказалось так и есть: яичники ужев 13-ом сегменте, а семенники ещев 10-ом!
Выберем для парных органов явно старые и явно новые функции, тогда, по основной контра связи, старые должны доминировать слева, новые – справа. Значит.
5. Обезьяны брать корм должны чаще левой рукой, а манипулировать (открывать шпингалет) – правой. Специальные полевые наблюдения и обширные эксперименты многих авторов это предсказание теории полностью подтверждают, но объяснить не могут (MacNeilage, 1987; Beck & Barton, 1972; Ettlinger, 1961, 1964; Milner, 1969; Gautrin, 1970; Itani, 1957, 1963; Tokuda, 1969, подробнее: Геодакян, 1993).
6. Поскольку “инструментальные” функции рук у человека явно моложе “неинструментальных”, то можно предсказать, что праворукость по первым д.б. выше, чем по вторым. Это так и есть: по этнографическим фильмам трёх традиционных (дописьменных) культур определяли эти величины и получили для первых – 84% ПР, а для вторых – 54% (т.к. у амбидекстров 50%, то, соответственно: 68% и 8%!) (Marchant et al., 1995).
7. Когда далекие предки человека переходили к прямохождению, новый центр управления возник в ЛП. Пока они овладевали хождением на двух ногах, ЛП решало связанные с ним проблемы: равновесия, сужения шейки аорты, создания клапанов в венах ног и т.д. Был период, когда двуногость была у мужчин уже генотипической, а у женщин еще фенотипической (за счет широкой нормы реакции, высокой обучаемости и конформности). Тогда, по новой теории, мужчины д.б. правоногими, а спустя много поколений, после транслокации, стать левоногими. Поэтому, согласно закону рекапитуляции, в онтогенезе современного человека, в определенном возрасте “детская правоногость” должна превращаться во ”взрослую левоногость”. Такой переход был открыт (Bonin, 1962). Было установлено, что у детей от 6 до 12 лет больше правая бедренная кость, т.е. они правоноги, а с 13 до 20 лет – левая! Такое парадоксальное явление никак иначе не объяснить. (Можно предсказать еще, что возраст транслокации в онтогенезе у девушек д.б. позже, чем у юношей).
8. В отличие от рук, имеющих много новых функций, у ног их мало. Этим объясняется, что человек, как вид праворук (~ на 90%) и левоног (Касинов, Никитюк, 1975). Если же тестировать руки по старой функции (хватательной), а ноги по новой (писать, рисовать), то теория предсказывает, что получим диаметрально противоположный вывод: человек, как вид окажется леворук (хватательный рефлекс, см. ниже п. 20) и правоног. Последнее легко проверить, закрыв глаза и представив, что пишете ногой по песку.
9. По той же логике, эволюционно старые звуки – средовые (шум дождя, ветра, моря, лай собаки, кашель) лучше должно улавливать левое ухо, а смысловые звуки (слова, числа) – правое. Эксперименты по дихотическому прослушиванию это предсказание полностью подтверждают (Blumstein et al., 1975; Curry, 1967; Harris, 1978).
10. По зрительному рецептору, по той же схеме, старые стимулы, вспышку света, лучше должны улавливаться в левом зрительном поле, тогда как новые – мелькание слов или чисел – в правом. Тахистоскопические эксперименты это также подтверждают (Спрингер, Дейч 1983).
11. Недавно калифорнийские ученые по той же методике предлагали испытуемым обычные слова (древнее) и брэнды (моложе). В полном согласии с теорией, лучшие результаты были по первым в левом зрительном поле, а по вторым – в правом.
12. То же самое и по тактильному рецептору: новые (незнакомые) предметы на ощупь лучше должны узнаваться правой рукой, а старые (хорошо знакомые) – левой. Это предсказание теории также подтверждают многочисленные эксперименты по дигаптическому узнаванию предметов (Спрингер, Дейч 1983; Witelson, 1976).
13. А по обонятельному рецептору, все д.б. наоборот, т.к у человека оно утрачивается, то чувствительнее д.б. левая ноздря. И это так: она, как правило, ỳже и чувствительнее, чем правая. Говорят потому, что она ỳже. Тогда я спрашиваю, почему она ỳже? Ответа опять нет.
14. В филогенезе, по теории, рукость менялась: амбидекстрия®праворукость® леворукость. Поскольку мужской пол – эволюционный авангард, то в этих группах должна расти доля мужчин. Этот прогноз теории тоже оправдывается. На каждую женщину приходится среди амбидекстров примерно 0,5, среди праворуких ~ 0,9, а леворуких ~ 5 мужчин (Briden, 1987).
15. Согласно новой теории, ПП – биологическое, видовое, ЛП – социокультурное, этническое, поэтому дисперсия (разнообразие в популяции) должна быть больше по вторым. Это полностью подтверждают результаты экспериментов на трех этнических группах в Америке: индейцев, негров и белых (1220 человек) (Спрингер, Дейч, 1983). А также на северных народах у нас (Аршавский, 1988).
16. Изоморфизм явлений дает огромные преимущества. Как в кроссворде, в котором перекрещены загадки пола и асимметрии: нахождение любого слова по горизонтали облегчает поиск вертикальных. Например, блестящее подтверждение главной идеи – дихронизма,что от матерей мы получаем старую генетическую информацию (о прошлом), а от отцов “последние новости” эволюции (о настоящем) – загадочное открытие двух групп английских ученых (Surani et al 1983,1984, McGrath et al. 1983,1984), показавших, что развитие внезародышевыхоболочек и плаценты, сугубо женских органов, определяют мужскиегены (а не женские), а развитие “общего” эмбриона, – женскиегены (а не М и Ж)! Ведь у эмбриона генов отца и матери поровну, а “среда” материнская (рис. 7). Единственная разница: эмбрион– эволюционно стараясистема, а оболочкии плацента – новые: они появились у высших млекопитающих.
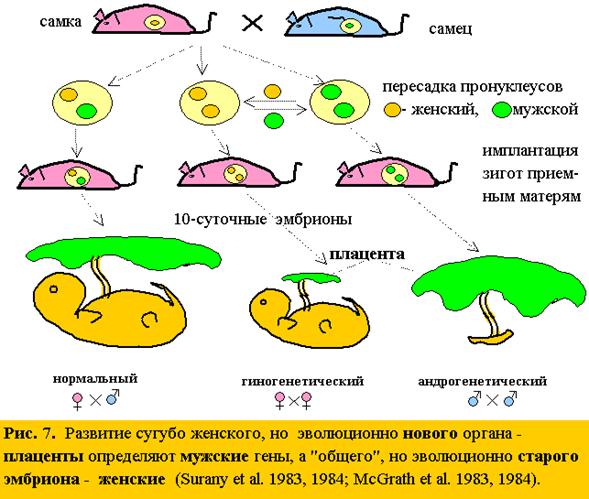
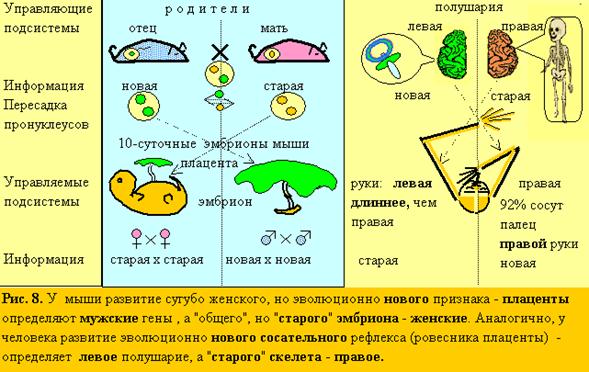
17. По той же логике (рис. 8), я попытался найти новыйповеденческий признак у эмбриона и предсказать его управление левым полушарием(аналогом отца), тогда, как эмбриогенезомв целом (морфология: череп, скелет), как старой системой, должно управлять правое полушарие(аналог матери). Новых признаков у эмбриона мало и ЛП (по теории – социальное) – почти пустое. И, тем не менее, есть – сосательный рефлекс (аналог и почти ровесник плаценты).Как известно, эмбрион в утробе матери сосет большой палец (см. 4.2.). Значит, этот палецд.б.правой руки, управляемой ЛП!А,управляемая ПП, левая сторона тела, д.б. крупнее, чем правая! Невероятно, но ультразвуковым методом было показано, что действительно в 92% случаев эмбрион сосет палец правойруки! (Hepper, 1991)! А у 3-х скелетов и черепов эмбрионов разного возраста в музее антропологии МГУ, левая сторона заметно больше правой. Тогда, как в музее института мозга, они, кажется, симметричны. Значит – эмбрионы были женского пола!
18. По той же причине степень “левополушарности“ (ЛП/ПП) д.б. минимальна для эмбриона и расти в онтогенезе. Это также соответствует действительности. Эмбриологи отмечали, что у многих видов левая сторона тела развивается обычно немного быстрее правой (Клименко, 1984). Установлен также ПП контроль внутриутробного развития человека (Walker, 1980).
19. Еще одно доказательство этого – соотношение длин левой и правой височных плоскостей: у младенцев оно равно 1,64, а у взрослых – 1,82 (Wada et al., 1975).
20. Безусловные рефлексы, как более старые, должны иметь “левый уклон” (ПП), а условные (молодые) – правый (ЛП). Хватательный рефлекс у новорожденных, несмотря на видовую праворукость человека, сильнее выражен у левой руки, при этом даже участие ладони сильнее слева, а пальцев – справа (MacNeilage, 1987).
21. Предсказанную, ЛП®ПП, транслокацию подтвердили при изучении выработки классического условного рефлекса, у человека, собак и кошек путем регистрации вызванных потенциалов и ЭЭГ. Авторы пишут: “…«закон Геодакяна» в равной мере справедлив и для фило, и для онтогенеза, и для процесса индивидуального обучения” (Симонов и др., 1995). Такое “расширение” теории – приятно, но пока, не очень понятно: нужно ли вводить третий масштаб времени.
22. Также подкрепляет и расширяет теорию открытие такой же транслокации в гипоталамусе (Павлова, 2001).
Таковы некоторые открытия и факты, которые существующие теории не могут объяснить, а новая концепция предсказывает и трактует естественно и просто.
Теория цис-транс организмов.
Гипотеза генотипической полушарности – фенотипической рукости.
По теории БСД (см. 3.1), две пары управляющих и управляемых органов (ЛП, ПП, ПР, ЛР), образуют два вида асимметрии: внутрипарную и межпарную,… Генотип — более конститутивное (фундаментальное) явление, чем фенотип, но мы… Информационные отношения полушарий мозга, как управляющих органов, и рук, как управляемых, аналогичны отношениям по…Истоки латеральной асимметрии.
Есть существенная разница между L-D и цис-транс изомерами. Первые по большинству физических и химических свойств неотличимы друг от друга, но резко… Известно, что у транс молекул, как более симметричных (упорядоченных), меньше…Морфологические новации, появившиеся в филогенезе человека при латеральной асимметризации и, связанные с ними, функциональные признаки в разных средах.
В филогенезе латеральная асимметрия появляется, у плацентарных млекопитающих. Поэтому, если, не вдаваясь в детали, сравнить схемы нейро-морфологических структур, опоссума(БС) и человека(ТА), то можно попытаться по разности судить о роли и механизмах латерализации. У БС предшественников человека существовали: 1. Генотипически обусловленная межпарная зеркальная асимметрия между симметричными управляющими органами – полушариями и симметричными управляемыми – “руками”. Это – главная асимметризация БС форм по оси нос–хвост. Её вектор показывает направление потока новой информации: перед®зад (см. разд. 4). 2. Простая схема контра-связей каждого полушария мозга с противоположной стороной тела. Смысл этого перекреста считается неясным (Walker, 1980).
Какие новые морфологические признаки, отсутствующие у опоссума, появились у человека и какие новые функциональные признаки они создают?
Морфологические признаки. 1. Мозолистое тело, состоящее из 200–350 млн. нервных волокон, соединяющих между собой соответствующие области коры двух полушарий мозга, кроме обонятельных и гипокампа. 2. Ипси связи – прямые нервные пути, соединяющие каждое полушарие с той же стороной тела (см. рис. 6).
Функциональные признаки. 1. Генотипическаявнутрипарная зеркальная асимметрия и специализация полушарий мозга,проявляющаяся в доминантности одного и субординатностидругого полушария по разным фазам эволюции функций. 2. Фенотипическаявнутрипарная зеркальная асимметрия рук (и других парных органов). 3. Поскольку внутрипарные (латеральные) асимметрии возникли на фоне межпарной асимметрии БС форм, то у ТА появляются 4 градации D-S (на рис.5, черная, темно-серая, светло-серая, белая). 4. Фенотипическая межпарная позиционная асимметрия организмов по цис-транс конфигурации доминантных полушария и руки (левши–правши). 5. Асимметрия кровоснабжения полушарий сонными артериями: D-рука повышает систолическое давление ипси-полушария, которое всегда у транс руких – S, у цис-руких – D, т.е. можно считать, что у цис руких, как бы, D-полушарие становится “сверхдоминантным”(Геодакян, 1993). 6. У них же, можно думать, соединение “сверх D” полушария через контра связь с S-рукой (максимально благоприятствует обучению S-руки новой функции). 7. Асимметрия численностисубпопуляций цис и транс особей (% левшей). 8. Асимметрия их дисперсий (разнообразия). 9. Асимметрия модальности субпопуляций (латерального диморфизма – отношения средних значений признака у левшей и правшей).
С другой стороны известно, что у высших млекопитающих главные эволюционные изменения, достигающие максимума у человека, наблюдаются в трех основных системах мозга: а) в ассоциативных структурах таламусаи коры, б) в области фронтальной коры, в) в системе мозолистого тела (Карамян, 1970). Если первые можно отнести к асимметрии по оси верх–низ, вторые – по оси перед–зад, то образование мозолистого тела, несомненно, относится к латеральной асимметрии, т.к. осуществляет межкортикальные связи двух полушарий.
Стало быть, все перечисленные выше эволюционные новации тесно связаны с латерализацией, но роль их неясна. Более того, считается парадоксальным, что в филогенезе специализация полушарий и развитие мозолистого тела происходили одновременно, т.к. думали, что межполушарные связи могут только нивелировать различия, а не создавать их, поэтому разные полушария меньше требуют связей, чем одинаковые (Прибрам, 1975). На самом деле все наоборот, т.к. только появление мозолистого тела, создает последовательное соединение полушарий (рис. 1) и дихронизм, т.е. асимметрию фаз эволюции.
Существующие теории оперируют только одной формой асимметрии – зеркально-внутрипарной асимметрией функцийи органови не учитывают межпарных отношений. Но, как мы видели, на уровне двух пар органов и выше (организма и популяции) она перестает “работать” и центральной фигурой становится межпарная цис-транс асимметрия.
Стало быть, новая концепция использует две “межпарные” идеи: 1. Гипотезу генотипической полушарности–фенотипической рукости (см. 6.1), которая вытекает из унаследованной от БС форм асимметрии перед–зад (рис. 3, 4). 2. Цис-транс рукость(асимметрия), которая более конститутивна, чем лево-право рукость. Это очевидно по частотам и дисперсиям (рис. 6).
6.5. Эволюционная роль мозолистого тела, контра и ипси связей.
Итак, у БС организмов была только контра связь, а у ТА – контра, мозолистое тело и ипсисвязи. Значит, первое более древнее образование, чем последние, по которым, согласно ЭТП, можно предсказать более высокую эволюционную продвинутость мужчин, т.е. бóльшее число нервных волокон в мозолистом теле и ипси связях, чем у женщин. Косвенным доказательством этого можно считать более асимметричные полушария и более высокие пространственно-зрительные способности у мужчин (Геодакян, 1987, 1993). Какова роль каждого типа связей?
Дихронная эволюция мозга требует последовательного соединения полушарий (рис. 1). Для этого возникло мозолистое тело, создающее зеркальную асимметрию полушарий: D и S-формы функций, полушарий, рук и других парных органов, но, только в транс позиции (т.к. наличие у БС форм только контра связи не позволяет создавать цис рукость, для этого нужны ипси связи). Тогда “цель” (назначение) ипси связей – создание цис конфигураций. Интересно отметить, что если мозолистое тело возникло раньше, чем ипси связи, то в этот период ЛРд.б. не ~10%, а ~1%! (Это предсказание теории для видов, имеющих мозолистое тело, но не имеющих ипси связей, если таковые существуют?). А что дает цис позиция? В чем её принципиальное отличие от транс позиции?
Таких отличий, по крайней мере три. 1. Более широкая фенотипическая дисперсия цис особей (чем транс), обеспечивающая их более тесную связь со средой (отбором); это значит, что у них, как и у мужского пола, ỳже норма реакции. 2. Повышенное артериальное давление D-полушария цис особей, которое можно интерпретировать, как “сверхдоминантность”, улучшающее поиск и, дающее возможность организму найти выход в экстремальной ситуации. 3. Уникальное соединение сверх-D полушария с S-рукой через контра связь, обеспечивающее максимальный D-S перепад, способствующий обучению рук новым функциям, т.к. для обучения требуется максимальный информационный потенциал, и широкий канал связи. Все эти особенности уникальны и присущи только цис рукости (см. рис. 6).
Управляющие подсистемы (полушария) связаны с управляемыми (сторонами тела) энергетическими (сонные артерии) и информационными (нервные пути) связями. Первые – только ипси, вторые и контра и ипси.
Можно выделить два контура регуляции: на уровне мономодальной, унитарной системы, сохраняющую её дисперсию (пример, популяция гомо-гетерозигот или субпопуляции одного пола, или одного полушария мозга, малые круги на рис. 11), и на уровне БСД, сохраняющие и регулирующие дисперсиии диморфизмы(большие круги). При этом контра пути осуществляют отрицательнуюобратную связь, которая сохраняет симметрию у первых и степень асимметрии у вторых (аналоги мужской Х-хромосомы), тогда как ипси пути и асимметричное кровоснабжение – положительнуюобратную связь (аналоги Y-хромосомы), меняющую равновесное состояние sи ПД (точно, как в холодильнике: отрицательная связь сохраняет заданную температуру, а положительная – меняет её) (Геодакян, 1993).
Эволюционная теория левшества – правшества (популяционная асимметрия).
Изоморфизм пол–латеральность позволяет по аналогии со вскрытым в ЭТП, “Экологическим правилом дифференциации полов” (Геодакян, 1991, 2000),… 7.1. Гипотеза эмбриональной гипоксии и “взрослой” гипероксии левого полушария.Заключение
Удалось ли мне показать правоту великого теоретика естествознания о плодотворности аналогий – судить читателям. Но больно и горько, что среди них не будет того, кто первым проверил и подтвердил экспериментально асинхронную асимметрию. По его инициативе писалась и эта статья. К 150-летию И.П.Павлова (кстати, ярко выраженного левши) главный редактор “Журнала ВНД” П.В.Симонов, сообщив, что юбилейными будут все номера 1999 г., попросил написать статью по асимметрии, сказав, – “она украсит наш журнал”. На мой скепсис, что биологи не любят умозрения, поразительно лаконично, со сдержанной улыбкой, по поводу эмпиризма, заметил – “чего-то прилил, чего-то отлил”. После таких “душеукрепляющих”, но обязывающих слов, я, естественно, бесконечно “улучшая”, статью так и не отдал.
Поэтому, поставив, наконец, точку, с признательностью посвящаю её светлой памяти академика Павла Васильевича Симонова,который, несмотря на большую занятость, всегда был доступен и находил время для научного общения (по субботам, в безлюдном институте).
СПИСОК ЛИТЕРАТУРЫ
Аршавский В.В. 1988. Межполушарная асимметрия в системе поисковой активности. Владивосток: ДВО АН СССР. 135 с.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М.: Наука. 1964. Т. 1. 432 с.
Бианки В.Л. Асимметрия мозга животных. Л.: Наука. 1985. 295 с.
Геодакян В.А. Роль полов в передаче и преобразовании генетической информации // Пробл. передачи информации. 1965а. Т. 1, № 1. С. 105-113.
Геодакян В.А. О существовании обратной связи, регулирующей соотношение полов // Проблемы кибернетики. 1965б. М.: Наука. Вып. 13. С. 187-195.
Геодакян В.А. О структуре эволюционирующих систем // Проблемы кибернетики. 1972. М.: Наука. Вып. 25. С. 81-91.
Геодакян В.А. Дифференциальная смертность и норма реакции мужского и женского пола. Онтогенетическая и филогенетическая пластичность // Журн. общ. биол. 1974. Т. 35. №3. С. 376-385.
Геодакян В.А. Концепция информации и живые системы // Журн. общ. биол. 1975. Т. 36. №3. С. 336-347.
Геодакян В.А. Асимметрия мозга и пол // Антропогенетика, антропология и спорт (Всес. симпоз.). Винница. 1980. Т. 2. С. 331-332.
Геодакян В.А. Системный подход и закономерности в биологии // Системные исследования Ежегодник 1984а. М: Наука. С. 329-338.
Геодакян В.А. Генетико-экологическая трактовка латерализации мозга и половых различий // Теория, методология и практика системных исследований (Всес. конф.). М. 1984б. С. 21-24.
Геодакян В.А. Системно-эволюционная трактовка асимметрии мозга // В кн.: Системные исследования. Ежегодник. М.: Наука. 1987. С. 355-376.
Геодакян В.А. Теория дифференциации полов в проблемах человека //В кн. Человек в системе наук. 1989. М: Наука. С. 171-189.
Геодакян В.А. Эволюционная трактовка асимметрии мозга // Человек, экология, симметрия (Межд. симпоз.). Минск. 1991. Навука i тэхника. С. 41-42.
Геодакян В.А. Эволюционная теория пола // Природа. 1991. №8. С. 60-69.
Геодакян В.А. Эволюционная логика функциональной асимметрии мозга // Докл. РАН. 1992. Т. 324. № 6. С. 1323-1326.
Геодакян В.А. Асинхронная асимметрия // Журн. высшей нервной деятельности. 1993. Т. 43, вып. 3. С. 543-561.
Геодакян В.А. Эволюционная роль половых хромосом // Генетика. 1998а. Т. 34. № 8. С. 1171-1184. (Russian J. of Genetics. 1998. V. 34. № 8. P. 986-998).
Геодакян В.А. Эволюция асимметрии, сексуальности и культуры (Что такое культура с точки зрения теоретической биологии) // Информационное мировоззрение и эстетика. (Тр. Межд. симп.) Таганрог. 1998б. С. 116-143.
Геодакян В.А. Эволюционные хромосомы и эволюционный половой диморфизм // Изв. РАН. Серия биол. 2000. № 2. С. 133-148. (Biol. Bulletin. 2000. V.27. №2. P. 99-113).
Геодакян В.А., Геодакян К.В. Новая концепция леворукоси // Докл. Акад. Наук. 1997. Т. 356. №6. С. 1-5. (Doklady Biol. Sci. 1997. V. 356. P. 450-454).
Денисова З.В. Механизмы эмоционального поведения ребенка. Л.: Наука. 1978. 143с.
Доброхотова Т.А., Брагина Н.Н. // Левши. М.: Книга. 1994. 231 с.
Иванов В.В. Чет и нечет: асимметрия мозга и знаковых систем. М.: Сов. Радио. 1978. 185 с.
Карамян А.И. Функциональная эволюция мозга позвоночных. Л.: Наука. 1970. 300с.
Касинов В.Б., Никитюк Б.А. Асимметрия // Большая мед. энциклопедия. М.: Советская энциклопедия. 1975. Т. 2. С. 257-258.
Кимура Д. Половые различия в организации мозга // В мире науки. 1992. №11-12. С. 355-376.
Клименко С.М. 1984. Симметрия // Большая мед. энциклопедия. М.: Сов. энциклопедия. Т. 23. С. 250-251.
Костандов Э.А. Асимметрия зрительного восприятия и межполушарное взаимодействие // Физиология человека. 1978. Т. 4. №1. С. 3-16.
Любищев А.А. Пробл. формы, систематики и эволюции организмов. 1982. М.: Наука. 196 с.
Малиновский А.А. Незавершенные идеи некоторых советских генетиков // Природа. 1970. №2. С. 79-83.
Павлова И.В. Функциональная асимметрия мозга при мотивационных и эмоциональных состояниях мозга. Автореф. докт. дисс. М. 2001. 35 с.
Прибрам К. 1975. Языки мозга. М.: Прогресс. 464 с.
Ратькин А.В., Андреев В.С. Окраска цветков – модель для изучения некоторых проблем генетики развития растенй // Усп. совр. генетики. М.: Наука. 1988. Вып. 15. С. 211-240.
Симонов П.В., Русалова М.Н., Преображенская Л.А., Ванециан Г.Л. Фактор новизны и асимметрия деятельности мозга // Журн. высшей нервной деятельности. 1995. Т. 45, вып. 1. С. 13-17.
Спрингер С., Дейч Г. Левый мозг, правый мозг. М.: Мир. 1983. 256 с.
Степанов Ю.М. Роль функциональной асимметрии мозга в регуляции психофизиологического состояния в условиях Крайнего Севера. Автореф. канд. дисс. Новосибирск. 1988. 16 с.
Томилин А.Г. Китообразные. М.: АН СССР. 1957. 756 с.
Annett M. Left, Right, Hand and Brain: The Right Shift Theory // 1985. Erlbaum Press, New Jersey.
Barton S.C., Surani M.A.H., Norris M.L. Role of Paternal and Maternal Genomes in Mouse Development / Nature. 1984. V. 311. P. 374-376.
Beck C.H.M. & Barton R.L. 1972. Deviation and Laterality of Hand Preference in Monkeys // Cortex. V. 8. P. 339-363.
Bishop, D.V.M. Handedness and Developmental Disorders // Blackwell Scientific Press. 1990. Oxford.
Blumstein S., Goodglass H.& Tartter V. 1975. The Reliability of Ear Advantage in Dichotic Listening // Brain and Language. V. 2. P. 226-236.
Brackenbridge C.J. 1981. Secular Variation in Handedness over ninety years // Neuropsychologia. V. 19. No 8. P. 459-461.2
Bonin G. von. 1962. Anatomical Asymmetries of the Cerebral Hemispheres // Interhemispheric Relation and Cerebral Dominance. Baltimore: Johns Hopkins Univ. Press.
Boklage C.E. Twinning, Nonrighthandedness, and Fusion Malformations: Evidence for Heritable Causal Elements Held in Common // Am. J. Med. Genetics. 1987. V. 28. P. 67-84.
Boucher J. Hand Preference in Autistic Children and their Parents // J. Autism & Childhood Schizophrenia. 1977. V. 7. P. 177-187.
Bryden M.P. & McManus I.C. Relations between Handedness and Immune Disorders // J. Clinical & Experimental Neuropsychology. 1992. V. 14.P. 89.
Bryden M.P. // Duality and Unity of the Brain. L.: Mackmillan. 1987. P. 55-70.
Bryden M.P. Laterality: Functional Asymmetry in the Intact Brain. Academic Press. 1982. NY.
Buffery A. & Gray J. Sex Differences in the Development of Spatial and Linguistic Skills // in: Gender Differences, their Ontogeny and Significance (Ed. C. Ounsted). 1972. Edinburg. P. 123-158.
Colby K.M. & Parkinson C. Handedness in Autistic Children // J. Autism & Childhood Schizophrenia. 1977. V. 7. P. 3-9.
Connolly K.J., Bishop D.V.M. The Measurment of Handedness: A Cross-cultural Comparison of Samples from England and Papua New Guinea // Neuropsychologia. 1992. V. 30. P. 13-26.
Coren S. Left-handedness and Accident Risk // Am. J. Public Health. 1991. V.79. P.1-2.
Coren S., Porac C. Fifty Centuries of Right-handedness: The Historical Record // Science. 1977. V. 198. P. 631-632.
Crews D. Animal Sexuality // Scientific American. Jan. 1994. P. 108-114.
Curry F.W.K. 1967. Comparison of Left-handed and Right-handed Subjects on Verbal and Nonverbal Dichotic Listening Tasks // Cortex. V. 3. P. 343- 352.
Daniel W.F. & Yeo R.A. Left-handedness and Accident Rates // Am. J. Public Health. 1991. V. 81. P. 1036-1037.
Daniel W.F., Yeo R.A. & Gangestad S.W. Left-handers Suffer More Head Injuries than do Right- handers // J. Clinical & Experimental Neuropsychology. 1992. V. 14.P. 77.
Ellis L. Left- and Mixed Handedness and Criminality: Explanations for a Probable Relationship // in Left-handedness and: Behavioral Implications and Anomalies. 1990. Elsevier, N.Y. P. 485-508.
Ettlinger G. 1961. Lateral Preferences in Monkeys // Behaviour. V. 17. P. 275-287.
Ettlinger G. & Moffett 1964 A. Lateral Preference in the Monkeys // Nature. V. 204. P. 606.
Gautrin D. & Ettlinger G. 1970. Lateral Preferences in Monkeys // Cortex. V. 6. P. 287-292.
Fleminger J.J., Dalton R. & Standag K.F. Age as a Factor in the Handedness of Adults // Neuropsychologya. 1977. V.15. P. 471-473.
Fraser F.C. Association of Neural Tube Defects and Parental Nonrighthandedness // Am. J. Hum. Gen. 1983. V. 35. P. 89.
Galaburda A.M. Asymmetry of Cerebral Neuroanatomy // in Biol. Asymmetry and Handedness (Ed. Bock & Marsh). 1991. Wiley, N.Y. P. 219-233.
Geodakian V.A. Sexual Dimorphism of Brain Asymmetry and Psychology // Towards a New Synthesis in Evol. Biology. Proc. Intern. Symp. Praha. 1987. P. 262-263.
Geschwind N.W. & Galaburda A.M. Cerebral Lateralization: Biol. Mechanisms, Associations, and pathology. MIT Press. 1987. Cambridge, MA.
Gilbert A.N. & Vysocky C.J. Hand Preference and Age in the US // Neuropsychologia. 1992. V. 30. P. 601-608.
Gualtiery T. & Hicks R.E. An Immunoreactive Theory of Selective Male Affliction // Behavioral & Brain Sci. 1985. V. 8. P. 427-441.
Habib M., Gayraud D., Oliva A., Regis J., Salamon G. & Khalh R. Effects of Handedness and Sex of the Morphology of the Corpus Callosum: A Study with Brain Resonance Imaging // Brain and Cognition. 1991. V. 16. P. 41-61.
Halpern D.F. & Coren S. Handedness and Life span // Eng. J. Med. 1991. V. 324. P. 998.
Harris, L.J. Sex Differences in Spatial Ability // Asymmetry of the Function of the Brain. (Ed. Kempbel). 1978. L.: Cambridge.
Heller W. 1993. Neuropsychological Mechanisms of Individual Differences in Emotion, Personality, and Arousal // Neuropsychology. V.7. No 4. P. 476-489.
Hepper P.G., Shahidullah S. & White R. Handedness in the Human Fetus // Neuropsychologia. 1991. V. 28. P. 1107-1111.
Itani J. 1957. Personality of Japanese Monkeys // Iden. V. 11. P. 29-33. (Jap).
Itani J., Tokuda K. Et al. 1963. The Social Contraction of Natural Troops of Japanese Monkeys in Takasakiyama // Primates. V. 4. P. 1-42.
Landauer A. Rate of Motor reaction in mtn and women //Percept. and Mot. Skills. 1981. V. 52. P. 90-97.
Lenneberg E.H. Biol. Foundation of Language. 1967. Wiley. N.Y.
Levy J. Lateral Differences in the Human Brain Cognition and Behav. Control // Cerebral Correlates of Conscious Experience. 1978. No 1.
Lishman W.A. & McMeekan E.L.R. Hand Preference Pattern in Psychiatric Patients // Brit. J. Psychiatry. 1976. V. 129. P. 158-166.
London W.P. Left-handedness and Alcoholism // in Left-handedness: Behav. Implications and Anomalies (Ed. Coren). 1990. Elsevier. N.Y.
Maccoby E.& Jacklin C. The Psychology of Sex Difference. 1974. Stanford.
MacNeilage P.F. The Evolution of Handedness in Primates // Duality and Unity of the Brain. 1987. London: Macmillan.
McGlone J. Sex Difference in the Human Brain Asymmetry: a Critical Survey // Behav. and Brain Sci. 1980. V. 3. No. 2. P. 215-263.
McGlone J. Sex Difference in Functional Brain Asymmetry // Cortex. 1978. V. 14. P. 122-128.
McGrath J., Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes // Cell. 1984. Vol. 37. P. 179-183.
McGrath J., Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes // Cell. 1984. Vol. 37. P. 179-183.
Marchant L.F., McGrew W.C., Eibl-Eibesfeldt I. Is Human Handedness Universal? Ethological Analyses from Three Traditional Cultures // Ethology. 1995. V. 101. 239-258.
Milner A.D. 1969. Distribution of Hand Preferences in Monkeys // Neuropsychology. V. 7. P. 375-377.
Nasrallah N.A., Keelor K. & McCalley-Winters M. Laterality Shift in Alcoholic Males // Biol. Psychiatry. 1983. V. 18. P. 1056-1067.
O’Callaghan M.J., Tudehope D., Dugdale A.E., Mohay H., Burns Y. Handedness in Children with Birthweights below 1000g. // Lancet. 1987. №1. P. 1155.
Porac C., Coren S. & Duncan P. Life Span Age Trends in Laterality // J. Gerontolody. 1980. V. 35. P. 715-721.
Porac C., Coren S. & Duncan P. Life Span Age Trends in Laterality // J. Gerontolody. 1980. V. 35. P. 715-721.
Rife D.C. Handedness, with Special Reference to Twins // Genetics. 1940. V. 25. P. 178-186.
Satz P., Orsini D.L., Saslow E. & Henry R. The Pathological Left-handedness Syndrome //Brain and and Cognition. 1985. V. 4. P. 27-46.
Smart J.L., Jeffrey C. & Richards B.A. A Retrospective Study of the Relationship between Birth History and Handedness at Six Years // Early Human Development. 1980. V.5. P. 79-88.
Surani M.A.H., Barton S.C., Norris M.L. Development of reconstituted mouse eggs suggest imprinting of the genome during gametogenesis // Nature. 1984. Vol. 308. P. 548-550.
Tokuda K. 1969. On the Handedness of Japanes Monkeys // Primates. V. 10. P. 41-46.5
Waber D. Sex Differences in Cognition: a Function of Maturation Rate // Science. 1976. V. 192. P. 572-573.
Walker S.F. Lateralization of Functions in the Vertebrat Brain // Brit. J. Psychol. 1980. V. 71. P. 329-367.
Wada J.A., Clark R. & Hamm A. Cerebral Hemisphere Asymmetry in Humans // Arch. Neurol. 1975. V. 32. P. 239-246.
Witelson S.F. Sex and the Single Hemisphere // Science. 1976. V. 193. P. 425-427.
Witelson S.F. Neuroanatomical Asymmetry in Left-handers: A Review and Implications for Functional Asymmetry // in: The Neuropsychology of Left-handedness (Ed. Herron). 1980. Acad. Press. N.Y.
Witelson S.F. The Brain Connection: The Corpus Callosum is Larger in Left-handers // Science. 1985. V. 229. P. 1223-1226.
Witelson S.F. Hand and Sex Differences in the Isthmus and Genu of the Human Corpus Callosum // Brain. 1989. V. 112. P. 799-835.
Yeo R.A. & Gangestad S.W. Developmental Origins of Variation in Human Hand Preference // Genetica. 1993. V. 89. P. 281-296.
Yeo R.A., Gangestad S.W. & Daniel W.F. Hand Preference and Developmental Instability // Psychobiology. 1993. V. 21. P. 161-168.
[1] Сокращения в тексте и рисунках: С, Ас – симметрия, асимметрия; М, Ж – мужчина, мужской, женщина, женский; D, S – доминантность, субординатность; ЛП, ПП, ЛР, ПР – левое, правое полушария, руки; ЛП, ПП, ПР, ЛР –лево, правополушарность, рукость (векторы); ШС, РС, БС – шаровая, радиальная, билатеральная симметрии, ТА – триаксиальная асимметрия; УС – унитарная система; БСД – бинарная сопряженная дифференциация; КП, ОП – консервативная, оперативная подсистемы БСД; ЭТП –эволюционная теория пола; ФАМ – функциональная асимметрия мозга; Д –диморфизм, ПД– половой, ЛД – латеральный, ЭПД –эволюционный, gsd – генотипический ПД; ДХ –дихронизм; БП –бесполые, ГФ –гермафродиты, РП –раздельнополые; Т – транс, Ц –цис; I – информация, DI – разность информации, Iп –информационный потенциал; СП, СЛ– соотношения, ДП,ДЛ– дисперсии полов и латеральности.ЛТр, ЛЦр, ПТр, ПЦр –лево (право) полушарная транс (цис) рукость.
– Конец работы –
Используемые теги: Эволюционные, Теории, асимметризации, организмов, мозга, тела0.09
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Эволюционные теории асимметризации организмов, мозга и тела
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.18 сек.
Новости и инфо для студентов