12) Полиплоидия у растений
ПОЛИПЛОИДИЯ ,эуплоидия –наследств. изменение, заключающееся в кратном увеличении числа наборов хромосом в клетках организма. Наиб, часто встречается у растений и простейших, а из многоклеточных животных — у дождевых червей. Возникает в результате нарушения расхождения хромосом в митозе или мейозе под действием высокой или низкой темп-ры, ионизирующих излучений, химич. веществ (как в природе, так и в эксперименте). При П. наблюдаются отклонения от диплоидного числа хромосом в соматич. клетках и от гаплоидного — в половых; могут возникать клетки, в к-рых каждая хромосома представлена трижды (Зп — триплоиды), четырежды (4п — тетраплоиды), пять раз (5п — пентаплоиды) и т. д. Различают автополиплоидию (кратное увеличение числа наборов хромосом одного вида), характерную, как правило, для видов с вегетативным способом размножения (автополиплоиды стерильны в связи с нарушением конъюгации гомологичных хромосом в процессе мейоза), и аллополиплоидию (изменение числа наборов хромосом на основе межвидовой гибридизации), при крой обычно происходит удвоение числа хромосом у бесплодного диплоидного гибрида, и он становится в результате этого плодовитым. П. имеет важное значение в эволюции культурных и дикорастущих растений (полагают, что около трети всех видов растений возникли за счет П.), а также нек рых групп животных (преим. партеногенетических). Полиплоиды часто характеризуются крупными размерами, повышенным содержанием ряда веществ, устойчивостью к неблагоприятным факторам внеш. среды и др. хозяйственно полезными признаками. Они представляют важный источник изменчивости и м. б. использованы как исходный материал для селекции (на основе П. созданы высокоурожайные сорта с.-х. растений, устойчивые к болезням). В широком смысле под термином «П.» понимают как кратное (эуплоидия), так и некратное (анеуплоидия) изменение числа хромосом в клетках организма.
13) Строение хромосом. Аутосомы и половые хромосомы. Гаплоидный и диплоидный набор хромосом.
Различают 4 типа строения хромосом: 1)телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); 2)акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом); 3)субметацентрические (с плечами неравной длины, напоминающие по форме букву L); 4)метацентрические (V-образные хромосомы, обладающие плечами равной длины). Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Аутосомы - парные хромосомы, одинаковые для мужских и женских организмов. В клетках тела человека 44 аутосомы (22 пары). Половые хромосомы - хромосомы, содержащие гены, определяющие половые признаки организма. В кариотипе (качественном и количественном наборе хромосом) женщин половые хромосомы одинаковые. В кариотипе мужчины - 1 одна крупная равноплечая половая хромосома, другая - маленькая палочковидная хромосома. Половые хромосомы женщин обозначают XX, а мужские половые хромосомы - XY. Женский организм формирует гаметы с одинаковыми половыми хромосомами (гомогаметный организм), а мужской организм формирует гаметы неодинаковые по половым хромосомам (X и Y).
У птиц, бабочек и некоторых видов рыб гомогаметен мужской пол. У петуха кариотип обозначается XX, а у курицы - XY.
Гаплоидный и диплоидный набор хромосом
Гаплоидный и диплоидный набор хромосом. Понятие плоидности определяется как количество хромосомных наборов в клетках (преимущественно) в ядрах. У живых организмов хромосомы могут быть непарными и парными. В клетках человека образуется диплоидный набор хромосом, то есть двойной. Такой набор структурно-функциональных элементов характерен для всех соматических клеток. Стоит отметить, что у каждого человека диплоидный набор хромосом состоит из 44-х аутосом и 2-х половых хромосом. Гаплоидный набор хромосом является одинарным набором непарных структурно-функциональных элементов половых клеток. Данный набор содержит 22 аутосомы и только 1 половую хромосому. Гаплоидный набор и диплоидный набор хромосом могут присутствовать в одно и то же время. Это происходит преимущественно при половом процессе. В этот момент гаплоидная и диплоидная фазы чередуются. С помощью деления полный набор образует одинарный. После этого два одинарных соединяются и образуют полный набор структурно-функциональных элементов и т.д.
Диплоидный набор хромосом является совокупностью хромосом, которая присуща всем соматическим клеткам. В ней все хромосомы, которые характерны для данного биологического вида, представлены в парах. У каждого человека диплоидный набор хромосом способен содержать 44 аутосомы и 2 половых структурно - функциональных элемента. Диплоидный набор хромосом характерен для зиготы и всех соматических клеток, кроме анэуплоидных, гаплоидных и полиплоидных клеток.
14) Моногибридное скрещивание. Схема скрещивания. Понятие о гомо- и гетерозиготности. Доминантность и рецессивность. Закон доминирования.
Моногибридное скрещивание
Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков. При этом скрещиваемые предки являются гетерозиготными по положению хромосомы в аллели.
Моногибридное наследование представляет собой пример наследования единственного признака (гена), различные формы которого называют аллелями. Например, при моногибридном скрещивании между двумя чистыми линиями растений, гомозиготных по соответствующим признакам — одного с жёлтыми семенами (доминантный признак), а другого с зелёными семенами (рецессивный признак), можно ожидать, что первое поколение будет только с жёлтыми семенами, потому что аллель жёлтых семян доминирует над аллелью зелёных. При моногибридном скрещивании сравнивают только один характерный признак.
Доминантный признак (А)- преобладающий признак, проявляющийся всегда в потомстве, в гомо и гетерозиготном состоянии. Рецессивный признак (а) -подавляемый признак, проявляющийся только в гомозиготном состоянии. Гомозигота -зигота, имеющая одинаковые аллели одного гена (АА, аа). Гетерозигота - зигота, имеющая противоположные аллели одного гена (Аа).
Моногибридное скрещивание по одной паре признаков. 1. При полном доминировании проявляется только доминантный признак.
2. При неполном доминировании признак имеет среднее (промежуточное) значение между
 Скрещивание гибридов
Скрещивание гибридов
 при полном доминировании.
при полном доминировании.
 при неполном доминировании.
при неполном доминировании.
 Закон доминирования: «При скрещивании двух гомозиготных организмов, отличающихся по альтернативным вариантам одного и того же признака, все потомство от такого скрещивания окажется единообразным и будет нести признак одного из родителей».
37)БИОГЕНЕТИЧЕСКИЙ ЗАКОН, одно из обобщений эволюционной биологии, связывающее индивидуальное развитие, или онтогенез, с историческим развитием, или филогенезом. Биогенетический закон, установленный немецкими учёными Ф. Мюллером (1864) и Э. Геккелем (1866), утверждает, что онтогенез всякого организма есть краткое повторение (рекапитуляция) основных этапов филогенеза вида, к которому данный организм принадлежит. Биогенетический закон находит множество подтверждений в данных сравнительной анатомии, эмбриологии и палеонтологии. Напр., у зародышей птиц и млекопитающих на определённой стадии эмбрионального развития появляются зачатки жаберного аппарата. Это объясняется тем, что наземные позвоночные произошли от дышавших жабрами рыбообразных предков. Опираясь на биогенетический закон и используя данные эмбриологии, можно воссоздавать ход исторического развития тех или иных групп организмов. Это особенно важно в тех случаях, когда для к.-л. группы неизвестны ископаемые остатки предковых форм, т. е. при неполноте палеонтологической летописи. В нач. 20 в. русский биолог А.Н. Северцев показал, что в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей. Филогенез же следует рассматривать как последовательный ряд генетически связанных онтогенезов.
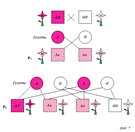
15) Анлизирующее скрещивание. Неполное доминирование. Схема скрещиваний. Анализирующее скрещивание- при полной доминантности среди особей с доминантными признаками невозможно отличить гомозиготы от гетерозигот, а в этом часто возникает необходимость (например, чтобы определить, чистородна или гибридна данная особь). С этой целью проводят анализирующее скрещивание, при котором исследуемая особь доминантными признаками скрещивается с рецессивной гомозиготой. Если потомство от такого скрещивания окажется однородным, значит, особь гомозиготна (ее генотип АА). Если же в потомстве будет 50% особей с доминантными признаками, а 50% с рецессивными, значит, особь гетерозиготна. Схему смотрите ниже.
Закон доминирования: «При скрещивании двух гомозиготных организмов, отличающихся по альтернативным вариантам одного и того же признака, все потомство от такого скрещивания окажется единообразным и будет нести признак одного из родителей».
37)БИОГЕНЕТИЧЕСКИЙ ЗАКОН, одно из обобщений эволюционной биологии, связывающее индивидуальное развитие, или онтогенез, с историческим развитием, или филогенезом. Биогенетический закон, установленный немецкими учёными Ф. Мюллером (1864) и Э. Геккелем (1866), утверждает, что онтогенез всякого организма есть краткое повторение (рекапитуляция) основных этапов филогенеза вида, к которому данный организм принадлежит. Биогенетический закон находит множество подтверждений в данных сравнительной анатомии, эмбриологии и палеонтологии. Напр., у зародышей птиц и млекопитающих на определённой стадии эмбрионального развития появляются зачатки жаберного аппарата. Это объясняется тем, что наземные позвоночные произошли от дышавших жабрами рыбообразных предков. Опираясь на биогенетический закон и используя данные эмбриологии, можно воссоздавать ход исторического развития тех или иных групп организмов. Это особенно важно в тех случаях, когда для к.-л. группы неизвестны ископаемые остатки предковых форм, т. е. при неполноте палеонтологической летописи. В нач. 20 в. русский биолог А.Н. Северцев показал, что в онтогенезе происходит повторение признаков не взрослых особей предков, а их зародышей. Филогенез же следует рассматривать как последовательный ряд генетически связанных онтогенезов.
15) Анлизирующее скрещивание. Неполное доминирование. Схема скрещиваний. Анализирующее скрещивание- при полной доминантности среди особей с доминантными признаками невозможно отличить гомозиготы от гетерозигот, а в этом часто возникает необходимость (например, чтобы определить, чистородна или гибридна данная особь). С этой целью проводят анализирующее скрещивание, при котором исследуемая особь доминантными признаками скрещивается с рецессивной гомозиготой. Если потомство от такого скрещивания окажется однородным, значит, особь гомозиготна (ее генотип АА). Если же в потомстве будет 50% особей с доминантными признаками, а 50% с рецессивными, значит, особь гетерозиготна. Схему смотрите ниже.
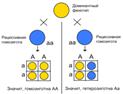 Неполное доминирование
Не всегда признаки можно четко разделить на доминантные и рецессивные. В этих случаях доминантный ген не до конца подавляет рецессивный ген из аллельной пары. При этом будут возникать промежуточные признаки, и признак у гомозиготных особей будет не таким, как у гетерозиготных. Это явление получило название неполного доминирования. Поясним его на примере.
При скрещивании растений ночной красавицы, имеющей пурпурные цветки (АА), с растением, имеющим белые цветки (аа), все растения первого поколения будут иметь промежуточную розовую окраску (рис. 1). Это не противоречит правилу единообразия гибридов первого поколения Г. Менделя: ведь действительно в первом поколении все цветки розовые. При скрещивании двух особей ночной красавицы из первого поколения во втором поколении происходит расщепление, но не в соотношении 3:1, а в соотношении 1:2:1, т.е. один цветок белый (аа), два розовых (Аа) и один пурпурный (АА).
Неполное доминирование
Не всегда признаки можно четко разделить на доминантные и рецессивные. В этих случаях доминантный ген не до конца подавляет рецессивный ген из аллельной пары. При этом будут возникать промежуточные признаки, и признак у гомозиготных особей будет не таким, как у гетерозиготных. Это явление получило название неполного доминирования. Поясним его на примере.
При скрещивании растений ночной красавицы, имеющей пурпурные цветки (АА), с растением, имеющим белые цветки (аа), все растения первого поколения будут иметь промежуточную розовую окраску (рис. 1). Это не противоречит правилу единообразия гибридов первого поколения Г. Менделя: ведь действительно в первом поколении все цветки розовые. При скрещивании двух особей ночной красавицы из первого поколения во втором поколении происходит расщепление, но не в соотношении 3:1, а в соотношении 1:2:1, т.е. один цветок белый (аа), два розовых (Аа) и один пурпурный (АА).
 46) Химический и ферментативный синтез генов вне организма.
Известны 2пути искусственного синтеза генов:
1.химический;
2.ферментативный.
Для химического синтеза необходимо иметь полностью расшифрованную последовательность нуклеотидов. Последовательность нуклеотидов в ДНК определяют по и-РНК. Впервые в 1970г. в США индийский ученый Корана осуществил искусственный синтез гена. Но этот ген не работал инвитро (в пробирке). Причиной являлся синтез только структурной части гена (в нем не было регуляторной части). В 1976г. был синтезирован ген, состоящий не только из структурного участка, но и регуляторных частей. Этот искусственный ген был введен в бактерию и функционировал в ней как природный. Химическим путем можно синтезировать небольшие по размеру гены прокариот. Синтез генов эукариот, состоящих из 1000 и более нуклеотидов путем химического синтеза создавать не удается. Кроме этого это метод очень трудоемкий и практически не применяется на практике. Наиболее успешным оказался ферментативный синтез. Это метод поколебал центральную догму молекулярной генетики, утверждающую, что считка информации происходит в направлении ДНК→и-РНК→белок. Оказалось, что РНК может быть предшественником ДНК. Подобное наблюдается у онкогенных РНК содержащих вирусов. С РНК вируса, попавшего в клетку, синтезируется ДНК-копия РНК с помощью фермента – обратная транскриптаза. Сам процесс называется обратная транскрипция(1970г.). На основе этих данных в 1972-1973г.г. во многих лабораториях мира были синтезированы гены кролика, мыши, утки, крысы. Но гены, синтезированные с помощью ревертаз (обратная транскриптаза) не имеют регуляторной части, а это препятствует функционированию искусственных генов в животных клетках, что ограничивает их использование.
Кроме того, и-РНК в клетках очень немного, и она не стойкая. В настоящее время рекомбинантные молекулы ДНК чаще всего получают путем гибридизации инвитро фрагментов ДНК вирусного и бактериального происхождения, и в меньшей степени эукариотического происхождения.
16) Закон расщепления. Генотип и фенотип. Понятие о гене как единице наследственности.
Закон расщепления, или второй закон Менделя.
Скрещиванием организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридное скрещивание. Закон расщепления: при моногибридном скрещивании во втором поколении гибридов наблюдается расщепление по фенотипу в соотношении 3:1 : около 3/4 гибридов второго поколения имеют доминантный признак, около 1/4 — рецессивный. Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несет доминантный признак, а часть — рецессивный, называется расщеплением. Следовательно, расщепление — это распределение доминантных и рецессивных признаков среди потомства в определенном числовом соотношении. Рецессивный признак у гибридов первого поколения не исчезает, а только подавляется и проявляется во втором гибридном поколении
Слово генотип имеет два смысла. В широком смысле - это совокупность всех генов данного организма. Но применительно к опытам того типа, которые ставил Мендель, словом генотип обозначают сочетание аллелей, которые контролируют данный признак (например, организмы могут иметь генотип AA, Aа или аа). Генотип — это совокупность всех генов организма, являющихся его наследственной основой. Таким образом, генотип - это:
- характерная для данного индивидуума вся совокупность генетических (геномных) характеристик и
- характеристика определенных пар аллелей , которые индивидуум имеет в исследуемом районе генома.
Фенотип — совокупность всех признаков и свойств организма, которые выявляются в процессе индивидуального развития в данных условиях и являются результатом взаимодействия генотипа с комплексом факторов внутренней и внешней среды. Термин "фенотип", как и генотип , используется в двух смыслах. В широком смысле - это совокупность всех признаков организма. Но применительно к моногибридному скрещиванию словом фенотип обычно обозначают тот признак, который в этом скрещивании изучается, например высокое растение имеет один фенотип, а карликовое - другой.
ГЕН - ЕДИНИЦА НАСЛЕДСТВЕННОСТИ.
46) Химический и ферментативный синтез генов вне организма.
Известны 2пути искусственного синтеза генов:
1.химический;
2.ферментативный.
Для химического синтеза необходимо иметь полностью расшифрованную последовательность нуклеотидов. Последовательность нуклеотидов в ДНК определяют по и-РНК. Впервые в 1970г. в США индийский ученый Корана осуществил искусственный синтез гена. Но этот ген не работал инвитро (в пробирке). Причиной являлся синтез только структурной части гена (в нем не было регуляторной части). В 1976г. был синтезирован ген, состоящий не только из структурного участка, но и регуляторных частей. Этот искусственный ген был введен в бактерию и функционировал в ней как природный. Химическим путем можно синтезировать небольшие по размеру гены прокариот. Синтез генов эукариот, состоящих из 1000 и более нуклеотидов путем химического синтеза создавать не удается. Кроме этого это метод очень трудоемкий и практически не применяется на практике. Наиболее успешным оказался ферментативный синтез. Это метод поколебал центральную догму молекулярной генетики, утверждающую, что считка информации происходит в направлении ДНК→и-РНК→белок. Оказалось, что РНК может быть предшественником ДНК. Подобное наблюдается у онкогенных РНК содержащих вирусов. С РНК вируса, попавшего в клетку, синтезируется ДНК-копия РНК с помощью фермента – обратная транскриптаза. Сам процесс называется обратная транскрипция(1970г.). На основе этих данных в 1972-1973г.г. во многих лабораториях мира были синтезированы гены кролика, мыши, утки, крысы. Но гены, синтезированные с помощью ревертаз (обратная транскриптаза) не имеют регуляторной части, а это препятствует функционированию искусственных генов в животных клетках, что ограничивает их использование.
Кроме того, и-РНК в клетках очень немного, и она не стойкая. В настоящее время рекомбинантные молекулы ДНК чаще всего получают путем гибридизации инвитро фрагментов ДНК вирусного и бактериального происхождения, и в меньшей степени эукариотического происхождения.
16) Закон расщепления. Генотип и фенотип. Понятие о гене как единице наследственности.
Закон расщепления, или второй закон Менделя.
Скрещиванием организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридное скрещивание. Закон расщепления: при моногибридном скрещивании во втором поколении гибридов наблюдается расщепление по фенотипу в соотношении 3:1 : около 3/4 гибридов второго поколения имеют доминантный признак, около 1/4 — рецессивный. Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несет доминантный признак, а часть — рецессивный, называется расщеплением. Следовательно, расщепление — это распределение доминантных и рецессивных признаков среди потомства в определенном числовом соотношении. Рецессивный признак у гибридов первого поколения не исчезает, а только подавляется и проявляется во втором гибридном поколении
Слово генотип имеет два смысла. В широком смысле - это совокупность всех генов данного организма. Но применительно к опытам того типа, которые ставил Мендель, словом генотип обозначают сочетание аллелей, которые контролируют данный признак (например, организмы могут иметь генотип AA, Aа или аа). Генотип — это совокупность всех генов организма, являющихся его наследственной основой. Таким образом, генотип - это:
- характерная для данного индивидуума вся совокупность генетических (геномных) характеристик и
- характеристика определенных пар аллелей , которые индивидуум имеет в исследуемом районе генома.
Фенотип — совокупность всех признаков и свойств организма, которые выявляются в процессе индивидуального развития в данных условиях и являются результатом взаимодействия генотипа с комплексом факторов внутренней и внешней среды. Термин "фенотип", как и генотип , используется в двух смыслах. В широком смысле - это совокупность всех признаков организма. Но применительно к моногибридному скрещиванию словом фенотип обычно обозначают тот признак, который в этом скрещивании изучается, например высокое растение имеет один фенотип, а карликовое - другой.
ГЕН - ЕДИНИЦА НАСЛЕДСТВЕННОСТИ.
| Единица наследственной информации получила название гена. Поскольку все свойства организма так или иначе определяются белками, то ген соответствует последовательности нуклеотидов, на базе которой синтезируется одна молекула белка. Ген неделим в функциональном отношении, т.е. все мутации одного гена изменяют один и тот же наследственный признак. Таким образом, достаточно протяженная молекула нуклеиновой кислоты, входящая в состав хромосомы, состоит из множества генов. Наследственные задатки, расположенные в пределах одной хромосомы, передаются потомкам более-менее сцеплено. Нарушения сцепления связаны с тем, что в процессе мейоза (редукционного деления) гомологичные хромосомы могут переплетаться и обмениваться участками (такой обмен носит название кроссинговера). Чем ближе в хромосоме располагаются гены, тем меньше вероятность обмена, поэтому сила сцепления позволяет оценить расстояние между генами и составить карту хромосомы. Уникальное свойство генов состоит в том, что они сочетают высокую устойчивость (неизменность в ряду поколений) со способностью к наследуемым изменениям - мутациям.
23) Типы хромосомного определения пола. Гомогаметньий, гетерогаметный пол. Кариотипы мужского и женского пола у разных видов.
Большинство живых организмов нашей планеты размножаются половым путем. Отсюда интерес, который проявили ученые к объяснению генетических основ определения пола и соотношения полов (мужского и женского) в природе. Было показано, что у насекомых, животных, человека и некоторых растений существуют специфические хромосомы, определяющие пол особи. Эти хромосомы получили название половых, а все остальные хромосомы - аутосом. При этом, у подавляющего числа организмов клетки женских особей имеют две одинаковые (морфологически) половые хромосомы, а клетки мужских особей разные, одна из которых полностью соответствует половым хромосомам женской особи. Естественно, что в результате в половые гаметы (яйцеклетки) женской особи попадают одинаковые половые хромосомы, а в мужские половые гаметы (сперматозоиды, спермин) - разные. Отсюда и определение у подобных организмов женского пола, как гомогаметный (гаметы по половым хромосомам похожи), а мужского, как гетерогаметный (гаметы несут разные половые хромосомы). Женские половые хромосомы были обозначены как X -хромосомы, а мужские как Y. Следовательно, женские гомогаметные особи несут две X хромосомы (XX), а мужские, являясь гетерогаметными, X и Y хромосомы (XY). Подобное хромосомное определение пола было установлено у человека, всех видов млекопитающих, дрозофилы и некоторых видов растений (конопля, дрема, щавель и др.). Наряду с этим оказалось, что у птиц, бабочек и некоторых растений (земляника) гомогаметным является мужской пол (ZZ), а гетерогаметным - женский (ZW). Третий тип определения пола был описан у кузнечика и многих видов полужесткокрылых (Hemiptera), прямокрылых (Orthotera). У них самки гомогаметны и несут две Х- хромосомы (XX), а вот самцы, несут только одну Х-хромосому (Х0). И наконец, у мхов в клетках женских особей присутствует одна Х-хромосома, а мужских - одна Y-хромосома. Во всех, вышеописанных случаях, женские хромосомы по размерам больше мужских. Большинство видов растений являются однодомными. У них в одном цветке образуются мужские и женские половые органы и одновременно формируются мужские и женские гаметы. И, независимо от вида опыления (самоопыление или перекрестное опыление), такие формы получили название гермафродитных форм. И наконец, интересно определение пола у пчел и муравьев. У этих насекомых нет половых хромосом. Самки - диплоидные особи и развиваются из оплодотворенных яиц, а самцы (трутни у пчел) из неоплодотворенных.
Гомогаметный пол— равногаметный пол, у которого половые хромосомы одинаковы. У большинства насекомых, млекопитающих, в том числе у человека, гомогаметный пол — женский (XX). У птиц, пресмыкающихся и бабочек гомогаметный пол — мужской (ZZ).
Гетерогаметный пол— разногаметный пол, у которого половина гамет имеет половую хромосому одного типа (X), половина — другого типа (Y). Так, у большинства насекомых, моллюсков, млекопитающих, в том числе и у человека, гетерогаметным является мужской пол; половина сперматозоидов несет Х-хромосому, половина — Y. Если в зиготе ХХ-хромосомы, то из нее разовьется особь женского пола, если XY - особь мужского пола. У птиц, пресмыкающихся, некоторых насекомых (бабочек) гетерогаметным является женский пол: половина яйцеклеток несет половую хромосому Z. половина — хромосому W. Если в зиготе находятся ZZ-хромосомы, то из нее будет развиваться особь мужского пола, а если ZW, то особь женского пола.
24) Балансовая теория определения пола. Интерсексуальность, фримартинизм, гинандроморфизм.
Изучая нерасхождение хромосом, К.Бриджес открыл важную роль баланса между числом наборов аутосом и числом Х-хромосом у дрозофилы в механизме определения пола. Оказалось, что при отношении числа Х-хромосом к числу наборов аутосом (Х/А) равном 1, развиваются самки.Если Х/А равно 0,5, то самцы образуются независимо от присутствия Y-хромосомы. Когда же отношение Х/А промежуточное между 0,5 и 1, насекомые приобретают черты интерсексуальности. Эта концепция получила название балансовой теории определения пола. У человека Х-хромосома направляет развитие организма в женскую сторону, а Y-хромосома в мужскую. При соотношении Х/Y равным 1 развивается нормальный мужчина, 2Х – нормальная женщина. Согласно балансовой теории определения пол особи определяется балансом генов, детерминирующих мужской и женский пол и локализованных в любых хромосомах генома. В настоящее время у человека описано 6 генов (3 в Х-хромосоме и 3 в Y-хромосоме), взаимодействие которых определяет пол особи. При отсутствии Y-хромосом и любом числе Х-хромосом особь определяется как женская. Балансовая теория определения пола показывает генетически обусловленную потенциальную бисексуальность всех раздельнополых организмов и их гамет.
Интерсексуальность — наличие у раздельнополого организма признаков обоих полов, причем эти признаки являются не полностью развитыми, промежуточными (ср. Гермафродитизм). Признаки обоих полов проявляются совместно на одних и тех же частях тела (ср. Гинандроморфизм). Эмбриональное развитие такого организма называется интерсексом, начинается нормально, но с определённого момента продолжается по типу другого пола. Чем раньше меняется направление развития организма, тем резче выражена у него интерсексуальность
Фримартинизм - уродство, нередко называемое неполным (ложным) гермафродитизмом, характеризующееся переразвитием клитора, принимающего форму мужского полового члена, при отсутствии или недоразвитии некоторых отделов женского полового аппарата. Псевдогермафродиты встречаются преимущественно при рождении разнополых двоен у коров, причем бычки, как правило, развиваются нормально и могут быть полноценными производителями, а телочки-фримартины бывают бесплодными. Фримартинизм регистрируется также у коз, свиней, реже у животных других видов.
Гинандроморфизм — аномалия развития организма, выражающаяся в том, что в одном организме крупные участки тела имеют генотип и признаки разных полов. Является результатом наличия в мужских и женских клетках организма наборов половых хромосом с разным количеством последних, как например у многих насекомых. Гинандроморфизм происходит как результат неправильного распределения половых хромосом по клеткам в ходе нарушенного созревания яйцеклетки, её оплодотворения или дробления. Особи — гинандроморфы наиболее ярко выражены у насекомых с четко проявляющимися признаками полового диморфизма, при этом морфологически выделяются следующие типы гинандроморфов:
· билатеральные, у которых одна продольная половина тела имеет признаки мужского пола, другая — женского;
· передне-задние, у которых передняя часть тела несет признаки одного пола, а задняя — другого;
· мозаичные, у которых перемежаются участки тела, несущие признаки разных полов.
30) Генетический код, его свойства: триплентность, неперекрываемость, вырожденность, универсальность. Биосинтез — синтез белка в клетке
Генетический код – система записи генетической информации в ДНК (РНК) в виде определенной последовательности нуклеотидов. Последовательность нуклеотидов определяет последовательность включения АК в синтезируемый белок. 3 нуклеотида – триплет – кодон – кодируют 1 АК. Совокупность триплетов и составляет генетический код.
Свойства генетического кода:
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии;
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислот, происходящий на рибосомах с участием молекул мРНК и тРНК. Процесс биосинтеза белка требует значительных затрат энергии.
Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и процессинг РНК, второй этап включает трансляцию. Во время транскрипции фермент РНК-полимераза синтезирует молекулу РНК, комплементарную последовательности соответствующего гена (участка ДНК). Терминатор в последовательности нуклеотидов ДНК определяет, в какой момент транскрипция прекратится. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, и редко происходит редактирование нуклеотидных последовательностей. После синтеза РНК на матрице ДНК происходит транспортировка молекул РНК в цитоплазму. В процессе трансляции информация, записанная в последовательности нуклеотидов переводится в последовательность остатков аминокислот.
Процессинг РНК
Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. К 5΄-концу присоединяется кэп, а к 3΄-концу поли-А хвост, который увеличивает длительность жизни иРНК. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК, — альтернативный сплайсинг.
Трансляция
У прокариот мРНК может считываться рибосомами в аминокислотную последовательность белков сразу после транскрипции, а у эукариот она транспортируется из ядра в цитоплазму, где находятся рибосомы. Скорость синтеза белков выше у прокариот и может достигать 20 аминокислот в секунду[1].
Процесс синтеза белка на основе молекулы мРНК называется трансляцией. Во время начальной стадии биосинтеза белков, инициации, обычно метиониновый кодон узнаётся малой субъединицей рибосомы, к которой при помощи белковых факторов инициации присоединена метиониновая транспортная РНК (тРНК). После узнавания стартового кодона к малой субъединице присоединяется большая субъединица и начинается вторая стадия трансляции — элонгация. При каждом движении рибосомы от 5' к 3' концу мРНК считывается один кодон путём образования водородных связей между тремя нуклеотидами (кодоном) мРНК и комплементарным ему антикодоном транспортной РНК, к которой присоединена соответствующая аминокислота. Синтез пептидной связи катализируется рибосомальной РНК (рРНК), образующей пептидилтрансферазный центр рибосомы. Рибосомальная РНК катализирует образование пептидной связи между последней аминокислотой растущего пептида и аминокислотой, присоединённой к тРНК, позиционируя атомы азота и углерода в положении, благоприятном для прохождения реакции. Ферменты аминоацил-тРНК-синтетазы присоединяют аминокислоты к их тРНК. Третья и последняя стадия трансляции, терминация, происходит при достижении рибосомой стоп-кодона, после чего белковые факторы терминации гидролизуют последнюю тРНК от белка, прекращая его синтез. Таким образом, в рибосомах белки всегда синтезируются от N- к C-концу.
31) Регуляция транскрипции у бактерий. Оперон.
Механизмы регуляции транскрипции у бактерий изучены достаточно хорошо. Эти примеры важны и для изучения биохимии животных, поскольку дают представление об общих молекулярных основах регуляции действия генов.
Рассмотрим пример индукции синтеза белков. Кишечная палочка в качестве пищевого вещества может использовать дисахарид лактозу. Превращения лактозы в клетках Е. coli начинаются с гидролиза при участии (З-галактозидазы (лактазы):При выращивании Е. coli на среде без лактозы р-галактозидаза в клетках обнаруживается в очень малых количествах. Если же в среду добавить лактозу, количество фермента увеличивается в сотни раз за несколько минут, т. е. происходит индукция синтеза фермента. Изучение молекулярных механизмов этого явления привело к созданию теории оперона. Опероном называют отрезок ДНК, содержащий структурные гены определенных белков и регуляторные участки. На рис. 4.22 приведена схема лак-тозного оперона.С участком оперона, называемым промотор, связывается РНК-полимераза. При ее движении по оперону происходит транскрипция структурных генов (3-га-лактозидазы, пермеазы, необходимой для транспорта лактозы в клетку, и галакто-зидтрансацетилазы. При этом получается одна молекула мРНК, содержащая матрицы для всех трех белков. Последующая трансляция этой мРНК ведет к образованию указанных белков. В результате транскрипции гена-регулятора образуется мРНК, служащая матрицей для синтеза белка-регулятора. Этот белок может присоединяться к оператору и тем самым блокировать транскрипцию структурных генов. Белок-регулятор может также соединяться с лактозой, при этом утрачивается его сродство к оператору. Если в среде есть лактоза, то белок-регулятор связан с ней, оператор свободен и возможна транскрипция — синтезируются белки, необходимые для усвоения лактозы. Если лактозы в среде нет, то белок-регулятор соединяется с оператором, транскрипции нет и белки, ненужные в этих условиях, не синтезируются. Биологическая целесообразность такой регуляции очевидна: достигается экономия веществ и энергии. Обязательным условием этого механизма регуляции является нестабильность мРНК: после исчерпания лактозы в среде мРНК должна быть быстро разрушена, чтобы прекратился синтез ставших ненужными белков. Это происходит путем гидролиза мРНК под действием РНКаз. Время полужизни мРНК в клетках бактерий измеряется немногими минутами.Примером регуляции путем репрессии синтеза может служить гистидиновый оперон бактерий Salmonella typhimurium. Этот оперон содержит 10 структурных генов, кодирующих 10 ферментов, необходимых для синтеза гистидина. Ферменты образуются только в том случае, когда в среде нет готового гистидина и клетки вынуждены сами синтезировать его из других веществ; добавление гистидина в среду прекращает синтез ферментов. Несмотря на противоположный результат индукции и репрессии синтеза белков, их молекулярные механизмы очень сходны. В действии гистидинового оперона легко разобраться, если на рис. 4.22 вместо «в присутствии лактозы» поставить «в отсутствие гистидина», вместо «в отсутствие лактозы» — «в присутствии гистидина» и вместо 3 структурных генов лак-тозного оперона — 10 структурных генов, кодирующих ферменты для синтеза гистидина.
40) Понятия о популяции и чистой линии. Методы изучения популяций По Н. В. Тимофееву-Ресовскому, популяция — это совокупность особей данного вида, в течение длительного времени (большого числа поколений) населяющая определенное пространство, состоящая из особей, могущих свободно скрещиваться друг с другом, и отделенная от таких же соседних совокупностей одной из форм изоляции (пространственной, сезонной, физиологической, генетической). Например, олени острова Колгуев изолированы от оленей, разводимых на материковой части Крайнего Севера, широкой полосой моря. В результате сформировалась особая популяция колгуевских оленей, отличающаяся от другой части этого вида генотипическими и фенотипическими признаками — они более крупные и обладают лучшей жизнеспособностью. В животноводстве под популяцией понимают группу животных одного вида, характеризующихся определенной численностью и ареалом распространения. Такая группа отличается от других популяций генетической структурой, экстерьерными, ин-терьерными и продуктивными качествами. Популяцией в животноводстве может быть отдельное стадо животных, порода или отродье. Обычно популяция — замкнутая группа. Ввоз в нее или вывоз из нее животных из других популяций ограничен, поэтому размножение в популяции осуществляется за счет подбора сам-цов и самок, принадлежащих к данной популяции. В Ярославской области, например, разводится популяция крупного рогатого скота ярославской породы. Каждая популяция характеризуется определенным генофондом, т. е. совокупностью аллелей, входящих в ее состав. Наряду с популяцией в генетике существует понятие «чистая линия* — это потомство, полученное только от одного родителя и имеющее с ним полное сходство по генотипу. Чистые линии могут быть созданы в растениеводстве у ; самоопыляющихся растений. В отличие от популяций они характеризуются полной гомозиготностью. Вследствие полной гомозиготности отбор в чистой линии невозможен, так как все особи, входящие в нее, имеют идентичный набор генов. Высокогомозиготных линейных мышей, крыс и других лабораторных животных создают в целях проведения различных экспериментов, например для проверки на мутагенность тех или иных препаратов, оценки вакцин и т. д. Популяция состоит из животных разных генотипов. Эффективность отбора в ней зависит от степени генетической изменчивости — соотношения доминантных и рецессивных генов.
34) Генные мутации.Генные мутации встречаются наиболее часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. Если под действием мутации изменяется один нуклеотид, говорят о точковых мутациях . Точковые мутации с заменой оснований разделяют на два класса: транзиции (замена пурина на пурин или пиримидина на пиримидин) и трансверсии (замена пурина на пиримидин или наоборот). Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций: сохранение смысла кодона ( синонимическая замена нуклеотида ), изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи ( миссенс-мутация ) или образование бессмысленного кодона с преждевременной терминацией ( нонсенс-мутация ). В генетическом коде имеются три бессмысленных кодона: амбер - UAG , охр - UAA и опал - UGA . В соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов (например амбер-мутация).
Генные Мутации, составляющие основную долю всех Мутации, вызывают чрезвычайно разнообразные изменения признаков организма, причём изменение одного гена обычно приводит к изменению нескольких признаков. Генные Мутации могут быть доминантными, полудоминантными и рецессивными. В результате Мутации ген может переходить в разные состояния (множественные аллели одного и того же гена), по-разному влияющие на контролируемые данным геном признаки организма. Мутантные гены могут отличаться от соответствующих нормальных тем, что специфический для данного гена продукт (чаще всего фермент) не образуется вовсе; образуется в меньшем или превышающем норму количестве; образуется продукт, инактивирующий или тормозящий продукт немутантного гена; вместо нормального образуется иной, не взаимодействующий с ним продукт, отсутствующий у немутантных особей. Претерпевший Мутации ген обычно столь же стабилен, как немутантный, из которого он произошёл; вследствие новой Мутации он может вернуться к исходному состоянию (обратные Мутации). Генные Мутации, как правило, вредны для организма, они нарушают жизненные процессы, протекающие в организме, снижают его жизнеспособность и плодовитость; нередко мутантный ген обусловливает гибель развивающегося организма (летальные Мутации). Реже возникают генные Мутации, сравнительно мало влияющие на жизнеспособность и плодовитость организма, ещё реже — улучшающие те или иные его свойства. Эта последняя категория генных Мутации, несмотря на свою относительную редкость, имеет огромное значение, т. к. даёт основной материал как для естественного отбора, так и для искусственного отбора, являясь необходимым условием эволюции и селекции.
. 35) Классификация хромосомных мутаций и их характеристика
Хромосомные мутации (аберрации) характеризуются изменением структуры отдельных хромосом. При них последовательность нуклеотидов в генах обычно не меняется, но изменение числа или положения генов при аберрациях может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма. Различают внутрихромосомные, межхромосомные и изохромосомные аберрации.
Внутрихромосомные аберрации — аберрации в пределах одной хромосомы. К ним относятся делеции, инверсии и дупликации.
Делеция — утрата одного из участков хромосомы (внутреннего или терминального), что может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития (например, делеция в регионе короткого плеча хромосомы 5, обозначаемая как 5р-, приводит к недоразвитию гортани, ВПР сердца, отставанию умственного развития). Этот симптомокомплекс обозначен как синдром кошачьего крика, поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье.
Инверсия — встраивание фрагмента хромосомы на прежнее место после поворота на 180°. В результате нарушается порядок расположения генов. Дупликация — удвоение (или умножение) какого-либо участка хромосомы (например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития). Межхромосомные аберрации — обмен фрагментами между негомологичными хромосомами. Они получили название транслокаций. Различают три варианта транслокаций: реципрокные (обмен фрагментами двух хромосом), нереципрокные (перенос фрагмента одной хромосомы на другую), робертсоновские (соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч, в результате образуется одна метацентри-ческая хромосома вместо двух акроцентрических).
Изохромосомные аберрации — образование одинаковых, но зеркальных фрагментов двух разных хромосом, содержащих одни и те же наборы генов. Это происходит в результате поперечного разрыва хроматид через центромеры (отсюда др. название — центрическое соединение).
38) Роль генетической информации на начальных стадиях онтогенеза. У животных в яйцеклетке до оплодотворения накапливается (в цитоплазме) большое количество рибонуклеиновых кислот всех трех типов: мРНК, рРНК и тРНК, — которые до оплодотворения находятся в неактивном состоянии. Они соединяются со специфическими белками-гистонами и образуют неактивные гранулы инфорсомы. Через несколько минут после оплодотворения часть молекул мРНК информосом освобождается от белка, поступает на рибосомы цитоплазмы яйцеклетки и начинает синтез определенных белков, необходимых %ля начального развития зиготы. Начальный период развития зиготы осуществляется под контролем генов материнского организма; мРНК яйцеклетки обеспечивает синтез белков до стадии поздней бластулы. С начала стадии гаструляции и в дальнейших процессах онтогенеза синтез белка осуществляется под контролем ядерных генов обеих родительских особей. В эмбриогенезе лягушки синтез мРНК возобновляется после 10 делений дробления, когда зародыш состоит приблизительно из тысячи клеток. А. А. Нейфах (1976) изучал первые стадии развития зародышей вьюна. Он обрабатывал оплодотворенные яйца лучами Рентгена. Дозы облучения были подобраны таким образом, чтобы прекратить деятельность ядра я нейтрализовать гены в молекулах ДНК. При облучении яиц сразу после оплодотворения или зародыша на стадии ранней бластулы развитие его шло нормально до поздней бластулы, а затем прекращалось, и наступала гибель. Следовательно, развитие эмбриона в этот период определяется иРНК, находящейся в цитоплазме клеток бластулы. Эта материнская иРНК и рРНК обеспечивают на ранней стадии дробления зиготы и бластулы синтез белков, необходимых для функционирования клеток и развития эмбриона. Развитие эмбриона прекращается на стадии гаструлы, так как для начала органогенеза нужны белки, синтез которых кодируется ядерными генами, локализованными в хромосомах материнской и отцовской особей. При облучении эмбрионов в период гаструляции, перед началом органогенеза и даже в период поздней бластулы развитие их прекращалось сразу после облучения. В некоторых случаях наблюдается наличие в цитоплазме яйцеклетки специальных фрагментов активной ДНК. Они синтезируют мРНК и кодируют синтез специфических белков в цитоплазме. У амфибий и рыб в цитоплазме в период созревания ооцитов и в яйцеклетках были обнаружены в большом количестве фрагменты молекул ДНК. В период созревания яйцеклеток происходит интенсивное насыщение цитоплазмы ДНК, мРНК, рРНК, тРНК, а также другими компонентами. У дрозофилы 15 фолликулярных клеток, окружающих яйцеклетку, проникают в нее цитоплазматическими выростами и насыщают цитоплазму митохондриями, ДНК, РНК, белками и другими компонентами. Таким образом, ядро зиготы, образовавшееся в результате слияния материнского и отцовского ядер и объединения их генетической информации, в начальный период развития зародыша не оказывает влияния на дробление зиготы и образование бластулы. Эмбрион в этот период развивается за счет РНК и других компонентов, находящихся в цитоплазме яйцеклетки. Критические периоды развития. Эмбриологи установили, что в онтогенезе, особенно на ранних стадиях развития, наблюдаются периоды, когда наиболее ярко выражена реакция эмбриона на воздействие внешних факторов. В эти периоды эмбрионы легко повреждаются, у них нарушаются процессы развития органов, что приводит к гибели эмбрионов либо к появлению уродств. Критические периоды обычно предшествуют началу соответствующего процесса органогенеза. В это время в соответствии с генетической программой развития особи усиливается синтез соответствующих белков, прекращается синтез предшествующих веществ, происходит перестройка обмена веществ в клетке. Критические периоды, как правило, наступают после поздней бластулы, когда дальнейшее развитие эмбриона осуществляется под контролем генетической информации обеих родительских особей. Наиболее изучены внешние факторы, влияющие в критические периоды на процесс онтогенеза у рыб, птиц, амфибий, рептилий, несколько меньше — у млекопитающих. У рыб нормальный онтогенез зависит от температуры воды и содержания в ней кислорода, причем у разных видов потребность в этих факторах различна: вьюн менее чувствителен к этим факторам, чем форель, лосось. У кур на эмбриогенез большое влияние оказывают температура и влажность воздуха в период инкубации. Эмбрионы особенно чувствительны к данным факторам на 2 — 3-й сутки инкубации, когда происходит образование системы кровообращения; на 8 — 9-е сутки — в период интенсивного морфогенеза — и на 19-е сутки, когда происходит переход зародыша к легочному типу дыхания. Критические периоды онтогенеза определены у хомяков, морских свинок, кроликов и других животных. У крупного рогатого скота наблюдается повышение эмбриональной смертности в первые дни развития зиготы, что свидетельствует о критическом периоде.
39) Регуляция синтеза белков у эукариот в процессе онтогенеза: стабильность мРНК, каскадная регуляция, гормональная регуляция
Биосинтез белка у эукариот.
1.Транскрипция – синтез и-РНК.
2.Посттранскрипционные процессы (процессинг) – образование зрелой и-РНК.
3.Трансляция – сборка полипептидной цепи.
4.Посттрансляционные процессы.
Синтез и-РНК происходит при участии фермента РНК-полимеразы, которая находит особый участок ДНК промотор, присоединяется к нему и, передвигаясь по цепи ДНК от 3´ к 5´концу, синтезирует по принципу комплементарности и
антипараллельности и-РНК. Дойдя до стоп-кодона, фермент прекращает свою работу. Так образуется незрелая и-РНК – первичный транскрипт. В результате процессинга выщепляются спейсеры и интроны, экзонные участки сшиваются – образуется зрелая и-РНК, на 5´конце которой расположен колпачок (кэп) с помощью которого и-РНК находит малую субъединицу рибосомы. С помощью лидера присоединяется к ней. За лидером расположен стартовый кодон (АУГ), затем кодоны (триплеты нуклеотидов), кодирующие аминокислоты, последним расположен стоп-кодон, за ним поли-адениловая последовательность из остатков адениловой кислоты (до 200 остатков).
Трансляция происходит в 3 этапа на рибосомах:
1.инициация (начало сборки белковой молекулы);
2.элонгация (наращивание цепи);
3.терминация (окончание сборки).
т-РНК доставляют аминокислоты к рибосомам. т-РНК имеет структуру, по форме напоминающую лист клевера. В т-РНК выделяют 3´конец, к которому присоединяется транспортируемая кислота. Средняя петля содержит антикодон -три нуклеотида, комплементарные кодону и-РНК, которые шифруют аминокислоту, транспортируемую данной т-РНК к рибосоме. Инициация – с помощью колпачка и-РНК находит малую субъединицу рибосомы, с помощью лидера присоединяется к ней. К ним присоединяется большая субъединица. В рибосоме выделяют два участка, связывающие т-РНК, аминоациальный (АЦ) и пептидильный (ПЦ) участки. В АЦ участке находится АУГ. К нему присоединяются т-РНК с аминокислотой – метианином. Происходит передвижка на один триплет. АУГ попадает в ПЦ участок, тогда как в АЦ участке располагается следующий за стартовым кодон.
Элонгация – циклически повторяющиеся события, при которых происходит узнавание антикодоном т-РНК кодона и-РНК, находящихся в АЦ участке, и их комплементарное взаимодействие. Между двумя аминокислотами, находящимися в АЦ и ПЦ участках, образуется пептидная связь, в результате предыдущая аминокислота теряет связь со своей
т-РНК и присоединяется к т-РНК, расположенной в АЦ участке. Т-РНК из ПЦ участка высвобождается и уходит в цитоплазму. Перемещение т-РНК, нагруженной пептидной цепочкой, из АЦ в ПЦ участок сопровождается продвижением рибосомы по и-РНК на один кодон. Так повторяется до тех пор, пока в АЦ не поступит стоп-кодон, для которого нет т-РНК. Сборка белка осуществляется со скоростью: у эукариот – 2 аминокислоты в секунду, у прокариот – 17 аминокислот в секунду.
| 44) Инбридинг, его биологические особенности и генетическая основа. Использование инбридинга в практике животноводства.
Инбридинг(от английского inbreeding: in - в, внутри, breeding - разведение) - один из методов разведения, при котором в пары сводят животных, происходящих от общих предков, то есть родственников.
О положительных и отрицательных сторонах инбридинга постоянно ведутся нескончаемые споры, и этот вопрос заслуживает особого рассмотрения. Противники этого метода указывают, что спаривание близких родственников влечёт за собой снижение плодовитости их потомства, общее ослабление телосложения потомков, большой процент мертворожденных и повышенную смертность от болезней, частое появление щенков, страдающих генетическими (наследственными) пороками и заболеваниями. Убеждение настолько сильно, что большинство собаководов на спаривание брата с сестрой среди породистых собак смотрят как на совершенно недопустимое и преступное явление. Сторонники этого метода напоминают, что все или почти все породы не только собак, но и других домашних животных были выведены с применением тесного инбридинга. Одновременно они указывают, что родственное спаривание позволяет закрепить в потомках желаемые качества исходных производителей.
Действительно, большинство популяций диких животных в природе возникает от одной пары или нескольких особей, случайно оказавшихся в новом, еще не заселенном данным видом месте. Совершенно очевидно, что даже если исходные производители были чужекровными, то через несколько поколений вся популяция будет состоять из родственников. Но регулятором отбора в данном случае будет сама
природа, которая допустит к продолжению рода самых приспособленных для данной местности. Некоторая часть лучшего в мире поголовья лошадей, коров, свиней, собак и других животных была так же получена путём инбридинга, который активно используется и в наше время для выведения более продуктивных пород в животноводстве. Родственное размножение является инструментом, способствующим переходу генофонда популяции в гомозиготное состояние. А факт ухудшения качества линий в процессе инбридинга лишь показывает, что немало аллелей, скрывавшихся в гетерозиготном состоянии у родоначальников линий, могли дать в гомозиготном состоянии отрицательный эффект. При этом повышается шанс внешнего проявления летальных и сублетальных генов, соответственно, рождения мертвых или неполноценных щенков. И частота таких случаев целиком зависит от частоты нежелательных аллелей в популяции. Итак, можно утверждать, что с помощью инбридинга в разведении животных решается целый ряд задач. Инбридинг на выдающихся производителей, сопровождаемый отбором, позволяет закрепить их ценные свойства в потомстве. Его используют для создания в породе четко различающихся линий или семейств. Инбридинг позволяет выявить генетические свойства особи и вести отбор против вредных рецессивных аллелей в популяции, способствуя улучшению качества поголовья. Но стоит особо подчеркнуть, что инбридинг - "инструмент" чрезвычайно серьёзный и требует от использующих его достаточных знаний и опыта, отсутствие
которых может нанести ощутимый вред породе.
48) Гибридизация соматических клеток и его значение. В основе метода лежит слияние клеток, в результате чего образуются гетерокарионы, содержащие ядра обоих родительских типов. Образовавшиеся гетерокарионы дают начало двум одноядерным гибридным клеткам. В 1965 английский ученый Г. Харрис впервые получил гетерокарионы, образованные клетками мыши и человека. Такую искусственную гибридизацию можно осуществлять между соматическими клетками, принадлежащими далеким в систематическом отношении организмам и даже между растительными и животными клетками. Гибридизация соматических клеток животных сыграла важную роль в исследовании механизмов реактивации генома покоющейся клетки и степени фенотипического проявления (экспрессивности) отдельных генов, клеточного деления, в картировании генов в хромосомах человека, в анализе причин злокачественного перерождения клеток. С помощью этого метода созданы гибридомы, используемые для получения моноклональных (однородных) антител.
Первый межвидовой гибрид при слиянии протопластов из клеток разных видов табака был получен в 1972 П. Карлсоном (США). Гибриды, полученные при слиянии протопластов, имеют важные отличия от половых гибридов поскольку несут цитоплазму обоих родителей. Возможно создание цибридов, наследующих ядерные гены одного из родителей наряду с цитоплазматическими генами обоих родителей. Особый интерес представляют цибриды растений, несущие цитоплазматические гены устойчивости к различным патогенам и стрессорным факторам от дикорастущих видов или цитоплазматические гены мужской стерильности. Слияние протопластов используют также для получения гибридов с ценными в хозяйственном отношении свойствами между отдаленными видами, которые плохо или вообще не скрещиваются обычным путем. Удалось, например, «ресинтезировать» рапс, являющийся естественным амфидиплоидом между турнепсом и капустой, получить соматический гибрид картофеля с томатами и т. д. При слиянии протопластов создают и новые клеточные линии-продуценты важных соединений.
причиной гетерозиса является то, что при скрещивании таких особей гаметы, из которых образуются зиготы, характеризуются достаточной … Кроме того, когда помесей получают от инбредных особей, скрещивание как бы… Большую роль в проявлении гетерозисаиграет и наследственная сочетаемость. Доказательством этому служит то хорошо …
вотных, имеющих в своем геноме чужой ген, принято называть трансгенными. Благодаря пере-носу генов у трансгенных животных возникают новые… ет закрепить их в потомстве и создавать трансгенные линии.
Так, были получены животные*биореакторы, способные выделять ценные биологические вещества. Например, в России…
В настоящее время в связи с установлением структуры молекул ДНК и их роли в передаче наследственной информации понятие гена претерпело дальнейшее… Размер гена определяется количеством парных оснований обеспечивающих…
· Летальная мутация, вызывающая нарушения в развитии хрящей у крыс, приводит к смерти за счет большого количества патологий в разных системах… · У овса окраска чешуйки и длина ости семени регулируются одним геном.
Летальные гены (также летали, более точно — летальные аллели) — гены, как правило, рецессивные, фенотипический эффект…
Сцепление генов и сцепленное наследование признаков. Группы сцепления. Кроссинговер как причина неполного сцепления генов. Одинарный и множественный кроссййговер.
Сцепленное наследование — феномен скоррелированного наследования определённых состояний генов, расположенных в одной хромосоме.
Полной корреляции не бывает из-за мейотического кроссинговера, так как сцепленные гены могут разойтись по разным гаметам. Кроссинговер наблюдается в виде расцепления у потомства тех аллелей генов и, соответственно, состояний признаков, которые были сцеплены у родителей.
изучая наследование разных пар признаков при дигибридном и полигибридном скрещиваниях, Морган и его сотрудники обнаружили большое число примеров сцепленного (совместного) их наследования. Все изученные признаки распределились на четыре группы сцепления в соответствии с числом и размерами хромосом у дрозофилы. На этом основании Морган сделал вывод о том, что гены, определяющие эти признаки, находятся в хромосомах. Гены, расположенные в одной хромосоме, представляют собой группу сцепления.
Сцепление генов — это совместное наследование генов, расположенных в одной и той же хромосоме.Количество групп сцепления соответствует гаплоидному числу хромосом. Например, у дрозофилы 4 группы сцепления, у человека 23, у крупного рогатого скота 30, у свиней 19 и т. д.
Мысль о расположении генов в хромосомах высказал Сеттон еще в 1902 г. Он обнаружил параллелизм в поведении хромосом в мейозе и наследовании признаков у одного из видов кузнечика. Дальнейшие исследования, проведенные Морганом, показали, что сцепление генов, расположенных в одной хромосоме, может быть полным или неполным.
одиночный кроссинговер – образуется только одна хиазма, что приводит к обмену только одним участком ДНК гомологичных хромосом. Это наиболее распространенный тип кроссинговера
двойной кроссинговер - образуются две хиазмы. Этот тип кроссинговера приводит к обмену двумя участками ДНК гомологичных хромосом
множественный кроссинговер - образуется более, чем две хиазмы между несестринскими хроматидами гомологичных хромосом. Далее они могут быть классифицированы как тройные (3 хиазмы), четвертные (4 хиазмы) и т.д
27)Строение молекул ДНК и РНК
Трансдукция – это перенос и рекомбинация генов у бактерий с помощью бактериофагов
Доказательства генетической роли нуклеиновых кислот
1. 1928 г. Опыты Фредерика Гриффита.
Признаки человека, наследование которых каким-то образом связано с полом, делятся на несколько категорий:
Признаки, ограниченные полом. Их развитие обусловлено генами, расположенными… Признаки, контролируемые полом или зависимые от пола. Развитие соматических признаков обусловено генами,…
Изменчивость. Классификация изменчивости. Классификация мутаций. Изменчивость
ИЗМЕНЧИВОСТЬ, способность живых организмов приобретать новые признаки и качества. Выражается в бесконечном разнообразии признаков и свойств у особей различной степени родства. При классификации типов и форм изменчивости подчеркиваются те или иные стороны этого универсального свойства. Учитывая причины и характер изменений, обычно выделяют два основных типа изменчивости.
Наследственная, или генотипическая, изменчивость обусловлена изменениями в генетическом материале (генотипе), которые передаются из поколения в поколение. Изменения в генотипе могут быть вызваны мутациями – изменениями в структуре генов и хромосом или изменениями числа хромосом в хромосомном наборе. При мутационной изменчивости возникают новые варианты (аллели) генов, причём мутации происходят сравнительно редко и внезапно. Другая форма генотипической изменчивости – т. н. комбинативная изменчивость, в основе которой лежит перекомбинация (перегруппировка) хромосом и их участков при половом размножении (в процессе мейоза и оплодотворения). В результате набор генов, а следовательно, и признаков у потомков всегда отличается от набора генов и признаков у родителей. Комбинативная изменчивость создаёт новые сочетания генов и обеспечивает как всё разнообразие организмов, так и неповторимую генетическую индивидуальность каждого из них.
Ненаследственная, или модификационная, изменчивость – способность организмов изменяться под действием различных факторов окружающей среды (температуры, влажности и т. п.). Этот тип изменчивости не связан с изменениями в генотипе и не наследуется. Однако пределы модификационной изменчивости любого признака – т. н. норма реакции – задаются генотипом. Степень варьирования признака, т. е. широта нормы реакции, зависит от значения признака: чем важнее признак, тем уже норма реакции. Модификационная изменчивость носит групповой характер – изменения (модификации) возникают у всех особей популяции, которая подвергается влиянию определённого внешнего воздействия. Другая её особенность – обратимость: обычно модификации сразу или постепенно исчезают при устранении вызвавшего их фактора.
Ч. Дарвин различал определённую (или групповую) и неопределённую (или индивидуальную) изменчивость, что по современной классификации совпадает соответственно с ненаследственной и наследственной изменчивостью. Следует помнить, однако, что это разделение в известной степени условно, т. к. пределы ненаследственной изменчивости определяются генотипом.
Типы мутаций. По характеру изменения генетического аппарата Мутации делят на геномные, хромосомные и генные, или точковые. Геномные Мутации заключаются в изменении числа хромосом в клетках организма. К ним относятся: полиплоидия — увеличение числа наборов хромосом, когда вместо обычных для диплоидных организмов 2 наборов хромосом их может быть 3, 4 и т. д.; гаплоидия — вместо 2 наборов хромосом имеется лишь один; анеуплоидия — одна или несколько пар гомологических хромосом отсутствуют (нуллисомия) или представлены не парой, а лишь одной хромосомой (моносомия) либо, напротив, 3 или более гомологичными партнёрами (трисомия, тетрасомия и т. д.). К хромосомным Мутации, или хромосомным перестройкам, относятся: инверсии — участок хромосомы перевёрнут на 180°, так что содержащиеся в нём гены расположены в обратном порядке по сравнению с нормальным; транслокации — обмен участками двух или более негомологичных хромосом; делеции — выпадение значительного участка хромосомы; нехватки (малые делеции) — выпадение небольшого участка хромосомы; дупликации — удвоение участка хромосомы; фрагментации — разрыв хромосомы на 2 части или более. Генные Мутации представляют собой стойкие изменения химического строения отдельных генов и, как правило, не отражаются на наблюдаемой в микроскоп морфологии хромосом. Известны также Мутации генов, локализованных не только в хромосомах, но и в некоторых самовоспроизводящихся органеллах цитоплазмы (например, в митохондриях, пластидах; см. Наследственность цитоплазматическая).
36) Геномные мутаций: эуплоидия (истинная полиплоидия), анеуплоидия (гетероплоидия) и их характеристика.
При геномных мутациях у организма-мутанта происходит внезапное изменение числа хромосом, кратное целому геному. Если через 2n обозначить число хромосом в исходном диплоидном геноме, то в результате геномной мутации, называемой полиплоидизацией , происходит образование полиплоидных организмов, геном которых представлен 4n, 6n и т.д. хромосомами. Различают аллополиплоидию , в результате которой происходит объединение при гибридизации целых неродственных геномов, и аутополиплоидию , для которой характерно адекватное увеличение числа хромосом собственного генома, кратное 2n.
Анеуплоидия -тероплоидия, явление, при к-ром клетки организма содержат изменённое число хромосом, не кратное гаплоидному набору. Отсутствие в хромосомном, наборе диплоида одной хромосомы наз. м оносомией, а двух гомологичных хромосом - нуллисомией; наличие дополнит, гомологичной хромосомы наз. трисомией. Организмы с такими изменениями числа хромосом наз. соответственно моносомиками, нуллисоми-ками и трисомиками. Осн. механизм возникновения А.- нерасхождение и потери отд. хромосом в митозе и мёйозе. Вследствие нарушения баланса хромосом А. приводит к понижению жизнеспособности и нередко к гибели анеуплоидов, особенно у животных (А. лежит в основе ряда хромосомных болезней). В генетич. анализе с помощью А. (скрещивая мутантов с анеуплоидами по определ. хромосомам) определяют, в какой группе сцепления находится исследуемый ген.
А. возникает в ходе деления клетки, когда обе хромосомы одной пары идут к одному полюсу, а к другому — ни одной; далее при оплодотворении первой гаметы другой гаметой с нормальным набором хромосом получается трисомик, а второй — моносомик. А. часто не совместима с жизнью или приводит к тяжелым патологиям. Напр., трисомия по 21-й паре хромосом у человека приводит к развитию синдрома Дауна.
Полиплоидия, эуплоидия (polyploidy, euploidy— наследственное изменение, заключающееся в кратном увеличении числа хромосом в клетках растений и животных; клетки могут содержать 3n (триплоиды), 4n (тетраплоиды), 5n (пентаплоиды) и т. д. Различают автополиплоидию (кратное увеличение числа хромосом одного вида) и аллополиплоидию (изменение набора хромосом на основе межвидовой гибридизации). Некратное изменение числа хромосом называют анеуплоидией (см. Анеуплоидия). П. возникает в результате слияния клеток или ошибок во время деления клетки. Некоторые ткани у животных (напр., склетные мышцы, гепатоциты, мегакариоциты, эпителий мочевого пузыря, миокард) часто полиплоидны в результате эндодупликаций генома или слияния клеток во время нормального развития. За счет П. клетки получают определенные преимущества, такие как устойчивость к повреждениям генома. Кроме того, увеличиваются размеры клеток, что предоставляет им большую гибкость и силу в ткани, подверженной механическим стрессам, напр., эпителий мочевого пузыря. Вместе с тем отклонение от нормальной плоидности в 2n может приводить к болезненным состояниям.
Харди и Вайнберг провели математический анализ распределения генов в больших популяциях, где нет отбора, мутаций и смешивания популяций. Они… Популяция находится в равновесии только тогда, когда в ней не происходит… генное равновесие и методы его определения
В исследованиях по генной инженерии часто используют кишечную палочку Е. coli. Геном этой бактерии представлен одной хромосомой (молекулой ДНК),…
Одна из важных задач - получение растений, устойчивых к вирусам, так как в настоящее время не существует других способов борьбы с вирусными… Еще одна задача связана с защитой растений от насекомых-вредителей.… Генные инженеры вывели трансгенные растения с удлиненным сроком созревания плодов. Такие помидоры, например, можно…
Возвратное скрещивание. Правило частоты гамет.
ВОЗВРАТНОЕ СКРЕЩИВАНИЕ (беккросс) - скрещивание гибрида (животных или растений) первого поколения с одной из родительских форм для выявления генотипа. Скрещивания между гибридной особью и одной из родительских форм называют Возвратными, Или Насыщающими, Скрещиваниями (беккроссами). Например, если гибрид Aа получен от скрещивания ААХаа, То скрещивания типа АаХАА Или АаХаа Будут возвратными. Такие скрещивания применяют, когда хотят усилить в гибриде проявление признаков какой-либо родительской формы. Особенно широко их используют в современной селекции при выведении сортов, устойчивых к болезням, создании стерильных аналогов и восстановителей фертильности. В подавляющем большинстве случаев результаты могут быть предсказаны заранее и повторно воспроизведены.
Мендель сформулировал правило чистоты гамет, состоящее в том, что у гетерозиготной особи наследственные задатки (гены) не перемешиваются друг с… Сущность правила (принципа) "чистоты гамет": 1) это гипотеза,…
Синтез белка в клетке
Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислот, происходящий на рибосомах с участием молекул мРНК и тРНК. Процесс биосинтеза белка требует значительных затрат энергии.
Введение
Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и процессинг РНК, второй этап включает трансляцию. Во время транскрипции фермент РНК-полимераза синтезирует молекулу РНК, комплементарную последовательности соответствующего гена (участка ДНК). Терминатор в последовательности нуклеотидов ДНК определяет, в какой момент транскрипция прекратится. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, и редко происходит редактирование нуклеотидных последовательностей. После синтеза РНК на матрице ДНК происходит транспортировка молекул РНК в цитоплазму. В процессе трансляции информация, записанная в последовательности нуклеотидов переводится в последовательность остатков аминокислот.
Процесс синтеза белка на основе молекулы мРНК называется трансляцией. Во время начальной стадии биосинтеза белков, инициации, обычно метиониновый…
Отбор и гибридизация являются основными и традиционными методами селекции растений. Применяя массовый или индивидуальный отбор, селекционер не… Для создания сортов растений с запрограммированными качествами ведется… Используя метод гибридизации с последующим отбором, селекционеры получили ценные высокоурожайные сорта пшеницы, ржи, …
Одомашнивание животных началось более 10 тыс. лет назад. Его центры в основном совпадают с центрами многообразия и происхождения культурных … К основным направлениям селекции животных относят:
— сочетание высокой продуктивности с приспособленностью пород к условиям среды конкретных природных зон; — повышение…
– Конец работы –
Используемые теги:
Строение, хромосом, аутосомы, Половые, хромосомы, Гаплоидный, диплоидный, набор, хромосом0.112
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ:
Строение хромосом. Аутосомы и половые хромосомы. Гаплоидный и диплоидный набор хромосом.
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
Еще рефераты, курсовые, дипломные работы на эту тему:
Строение хромосом. Аутосомы и половые хромосомы. Гаплоидный и диплоидный набор хромосом.
Инбридинг от английского inbreeding in в внутри breeding разведение один из методов разведения при котором в пары сводят животных... О положительных и отрицательных сторонах инбридинга постоянно ведутся... Действительно большинство популяций диких животных в природе возникает от одной пары или нескольких особей случайно...
Набор хромосом: соматические клетки имеют диплоидный, половые клетки – гаплоидный набор хромосом
Эмбриогенезу человека предшествует прогенез процессы развития половых клеток т е овогенез и сперматогенез Первые стадии развития половых... В результате гаметогенеза образуются половые мужские женские клетки которые... Яйцеклетка человека имеет диаметр около мкм окружена первичной оолеммa или собственная оболочка и вторичной...
Теория химического строения органических соединений. Электронная природа химических связей. Предпосылки теории строения. Теория химического строения. Изомерия
Органические вещества в своем составе наряду с другими элементами всегда содержат углерод. Изучение соединений углерода — их строения, химических… Из всех химических элементов только углерод образует такое большое число… По образованию оксида углерода (IУ) при горении или по обугливанию вещества при нагревании легко установить…
По идее Максвелла изменяющееся магн. поле является порождением вихревого эл. поля, а это поле создаёт индукционный ток
Если рассматривать возникновение ЭДС индукции в движущемся проводнике в этом случае ЭДС возникает благодаря силе Лоренца Если проводник неподвижен... По идее Максвелла изменяющееся магн поле является порождением вихревого эл... Вынужденные колебания ДУ вынужденных колебаний и его решение...
Половые хромосомы и аутосомы. Генотип
Взаимодействуют между собой как аллельные, так и неаллель-ные гены. Примером взаимодействия аллельных генов является неполное доминирование, т. е.… Так, скрещивание двух растений ночной красавицы - одного с красными, а другого… Это явление называется плейотропией. Например, у человека есть ген, который отвечает одновременно за развитие дефекта…
Экранирование электрического поля в полупроводниках. Дебаевская длина экранирования. Эффект поля
Система с зарядами экранирует внешнее поле если носители заряда связаны с атомами то уравнение Пуассона описывает экранирование электрического... уравнение Пуассона описывает экранирование эл поля в среде...
Наследственность. Классификация генов. Набор хромосом
Наследственность свойство живых организмов обеспечивать структурную и функциональную преемственность между поколениями а также специфический... Для того чтобы полноценно определить роль наследственности необходимо четко... Структурную и функциональную организацию ДНК...
На початку речення перед приголосними: В процессе разработки шахтного поля / У процесі розробки шахтного поля
НАУКОВО ТЕХНІЧНИХ ТЕКСТІВ... В основі вивчення особливостей наукового стилю української літературної мови у вищих навчальних закладах ВНЗ лежить...
МАГНИТНОЕ ПОЛЕ. ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ПОЛЯ
МАГНИТНОЕ ПОЛЕ ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ПОЛЯ...
Электрический заряд. Электрическое поле. Поле точечного заряда
На сайте allrefs.net читайте: " Электрический заряд. Электрическое поле. Поле точечного заряда"
0.03
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Наша политика приватности обеспечивает 100% безопасность и анонимность Ваших E-Mail
|






Новости и инфо для студентов