рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- АНТРОПОЛОГИЯ
Реферат Курсовая Конспект
АНТРОПОЛОГИЯ
АНТРОПОЛОГИЯ - раздел Философия, Высшее Образованиие ...
Высшее образованиие
 А. Е. Хомутов
А. Е. Хомутов
АНТРОПОЛОГИЯ
Учебное пособие
Издание третье
РОСТОВ-НА-ДОНУ
Фени
ББК 28.6 Х76
Рецензенты:
Зав. каф. нормальной физиологии НГМА, доктор биологических наук, проф. И.В. Мухина
Зав. каф. анатомии и физиологии НГПУ, доктор медицинских наук, проф. В.И. Щербаков
Научный редактор:
КульбаСН., К.6.В., доцент кафедры
физиологии человека и животных
Ростовского госуниверситета
Хомутов А. Б.
X 76 Антропология. Ростов н/Д: «Феникс», изд. 3-е, 2004. (Серия «Высшее образование») — 384 с.
В пособии изложены материалы по трем основным разделам антропологии: морфология человека, антропогенез и этническая антропология, или расоведение. Учебное пособие рекомендовано студентам психологических и биологических факультетов, преподавателям вузов и аспирантам, всем интересующимся проблемами становления человечества.
ББК 28.6
ISBN 5-222-05286-9
© Хомутов А. Е., 2004
© Оформление: изд-во «Феникс», 2004
ВВЕДЕНИЕ
Термин «антропология» имеет греческое происхождение и означает дословно «наука о человеке» (антропос — человек; логос — наука). В отечественной науке принято строгое разделение терминов «антропология», «этногра-фвя», «археология». Под археологией понимают науку, изучающую историческое прошлое человека по вещественным источникам, под этнографией — отрасль истории, исследующую все стороны культуры и быта ныне живущих народов, происхождение этих народов, историю их расселения, передвижения и культурно-исторических взаимоотношений. Антропология — отрасль естествознания, которая изучает происхождение и эволюцию физической организации человека и его рас. Но так как человек представляет собой существо качественно своеобразное, жизнь которого может протекать только в условиях общества и коллективно осуществляемого производства, то ясно, что антропологию нельзя поставить в один ряд с частными разделами зоологии. Антропология не может изучать человека с тех же позиций, с каких энтомология изучает насекомых, орнитология — птиц. Основной задачей антропологии является изучение процесса перехода от биологических закономерностей, которым подчинялось суще-
ствование животного предка человека, к закономерностям социальным. Таким образом, антропология занимает в кругу биологических наук особое место. Имея предметом своего исследования человека, она не может не выйти за пределы естественно-исторических вопросов и вступает в ту область знаний, где действуют факторы социально-исторические. Из этого пограничного положения антропологии в ряде наук и вытекает ее отношение к смежным отраслям знания. Антропология неразрывно связана с другими биологическими науками и вместе с тем тесным образом соприкасается с науками общественными.
Антропология включает в себя три основных раздела:
1) морфологию; " 2) антропогенез;
3) расоведение, или этническую антропологию. Раздел морфологии разрешает вопросы, связанные:
а) с индивидуальной изменчивостью физического типа;
б) с его возрастными изменениями от ранних стадий
зародышевого развития до старости включительно;
в) с явлениями полового диморфизма;
г) с анализом тех особенностей физической организа
ции человека, которые возникают под влиянием раз
личных условий жизни и труда.
Раздел морфологии состоит из:
1) мерологии (от греческого «мерос» — часть), и
щей вариации отдельных органов человека,
ных тканей, а также их взаимную связь;
2) соматологии (от греческого «сома» — тело), которая
изучает строение человеческого тела в целом, т.е.
закономерности вариаций роста, массы, окружности
груди, пропорций и т.д.
Важным подразделом соматологии является та отрасль антропологии, которая ставит своей задачей уста^ новление стандартов или норм размеров человеческого
тела и разрабатывает методы расчета, позволяющие установить, как часто встречаются те или иные отклонения от этих комбинаций. Большое значение имеет морфология для установления норм физического развития в разных возрастах, а также вариаций телосложения и их связи с физиологическими особенностями организма.
Наиболее важными вспомогательными дисциплинами для раздела морфологии являются нормальная анатомия, эмбриология и гистология человека. Необходимо отметить различия между нормальной анатомией и морфологией. Нормальная анатомия изучает человека как некоторый обобщенный тип и дает суммарную характеристику «среднего» человека. Морфология сосредотачивает свое внимание на вариациях типа и стремится постигнуть причины, закономерности и значение этих отклонений.
Раздел антропогенеза концентрирует свое внимание на тех изменениях, которые претерпевает природа ближайшего предка человека, а затем и самого человека в течение четвертичного периода. Это морфология'человека и его предшественников, рассматриваемая во времени, измеряемом геологическим масштабом. В разделе антропогенеза рассматриваются вопросы о месте человека в сис- теме животного мира, отношение его как зоологического вида к другим приматам, восстановления пути, по которому шло развитие высших приматов, выделения стадий в процессе человеческой эволюции, изучения условий и причин становления человека современного вида.
Раздел антропогенеза включает:
1) приматоведение, т.е. изучение современных и иско
паемых обезьян и полуобезьян;
2) эволюционную анатомию человека;
3) палеоантропологию, т.е. изучение ископаемых форм
человека.
Необходимыми вспомогательными дисциплинами для этого раздела являются: из наук естественно-истори-
ческих — геология четвертичного и третичного периодов, физиология высшей нервной деятельности; из наук социально-исторических — археология палеолита; из наук философских — психология.Расоведение, или этническая антропология, изучает классификацию рас, закономерности изменений расовых типов, распространение их по территории Земли, "причины расообразования. В числе смежных дисциплин, с которыми особенно тесно соприкасается расоведение, следует назвать из круга биологических наук — генетику и биометрию, а из наук социально-исторических — археологию позднего палеолита и последующих эпох, этнографию, языковедение и историю.
МОРФОЛОГИЯ ЧЕЛОВЕКА
Глава1. АНТРОПОМЕТРИЯ
 В основе антропологических методов исследования лежит антропометрия, или измерение размеров человеческого тела. По тому объекту, который служит предметом измерения, различают соматометрию — измерение живого человека, остеометрию — измерение костей скелета и краниометрию— измерение черепа.
В основе антропологических методов исследования лежит антропометрия, или измерение размеров человеческого тела. По тому объекту, который служит предметом измерения, различают соматометрию — измерение живого человека, остеометрию — измерение костей скелета и краниометрию— измерение черепа.
Одним из основных требований при проведении антропометрических исследований является их строгая унификация. Лишь единообразие приемов, использование точного, предварительно выверенного инструментария могут обеспечить достоверность полученных результатов. При массовых измерениях необходима одновременная работа нескольких исследователей, при этом каждый из них работает с одним инструментом, а один из исследователей фиксирует результаты.
При массовых обследованиях целесообразно для раздевания и измерений использовать отдельные смежные помещения, в которых поддерживается постоянная комнатная температура. Пол должен быть строго горизонтальным, его следует покрыть ковриком и одноразовой салфеткой, чтобы испытуемый мог стоять босыми ногами.
Измерения должны проводиться в первой половине дня натощак или через 2—3 часа после еды, в одно и то же время суток, так как длина тела к концу дня уменьшается на 1—2 см в связи с уплощением сводов стопы, межпозвонковых хрящей, снижением тонуса мускулатуры, а вес увеличивается в среднем почти на 1 кг. При измерении в более позднее время испытуемому рекомендуется полежать 10—15 минут.
При измерениях обследуемый находится в позе, близкой к положению тела, принимаемому по команде «смирно»: пятки вместе, носки врозь, ноги выпрямлены и прижаты друг к другу, живот подобран, руки опущены вдоль туловища, пальцы выпрямлены и прижаты друг к другу. Голова фиксируется так, чтобы козелок ушной раковины и нижний край глазницы находились в одной горизонтальной плоскости.
Длина тела (рост стоя) измеряется с помощью станкового ростомера или металлического антропометра.
Станковый ростомер представляет собой стойку длиной 2 м, укрепленную на широкой прочной платформе. По стойке передвигается муфта с планшеткой. На боковых поверхностях стойки имеются сантиметровые деления; с одной стороны отсчет ведется от платформы, с другой — от поверхности скамьи. На площадке ростомера укреплена откидная скамья, используемая для измерения роста сидя. Высота скамьи при измерении детей 25 см, взрослых — 35—40 см (рис. 1).
Станковым ростомером можно измерить рост только стоя и сидя, а антропометром — и различные длин-ники: длину всей части верхней или нижней конечности, длину туловища и т. д. Антропометр портативен и удобен в экспедиционных условиях.

|
| Рис. 1. Ростомер |
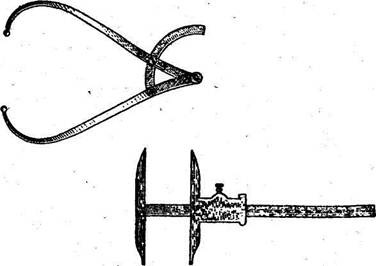
Антропометр (рис. 2) состоит из четырех полых равной длины металлических штанг, плотно вставляемых одна в другую в соответствии с цифровыми указателями, обозначенными на концах. Общая длина антропометра 2 м. На конце верхней штанги неподвижно укреплена муфта, в которую
|
Рнс.2. Металлический штанговый антропометр Мартина в разобранном виде
может вставляться линейка. Вторая муфта свободно перемещается по всей длине антропометра; она имеет вырез, через который видны деления. В эту муфту вставляется линейка скошенным концом кверху. По всей длине антропометра размещается шкала с ценой деления 1 мм, отсчет ведется снизу вверх.
Измерения поперечных и глубинных размеров тела производятся толстотным циркулем или верхней штангой антропометра. Для измерения диаметров применяют толстотные циркули (большие и малые) (рис. 3).
Толстотный циркуль состоит из двух ножек, скрепленных дугой или линейкой с делениями. По этой шкале ведется отсчет во время фиксации циркуля в установленном положении при измерениях. Циркуль толстотный малый (краниометр) устроен аналогично большому и от личается лишь меньшими размерами.
Обхватные размеры тела человека (окружности) измеряют сантиметровой лентой. Желательно, чтобы такая лента имела длину 1,5 м и была изготовлена из малорастяжимого материала.
Рве. 3. Толстотный н скользящий циркули
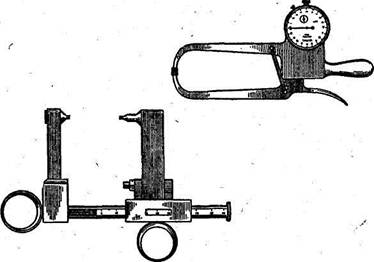
Определение толщины подкожного жирового слоя производится с помощью калипера — прибора, который позволяет производить измерение при стандартном давлении (10 г/мм2) и площади каждой измерительной ножки (90 мм2) или скользящего штангового циркуля (рис. 4).
Вес тела определяют на специальных медицинских весах, которые обеспечивают высокую надежность и точность измерений.
Мышечная сила измеряется динамометрами различной конструкции (рис. 5). Для определения силы кисти обычно пользуются динамометром Коллена. Отдельно измеряется сила правой и левой кисти. Силу экстензоров спины (разгибателей туловища) измеряют с помощью так называемого станового динамометра (рис. 6).
Антропометрические измерения с помощью антропометра и циркулей производятся с точностью до 1 мм. Измерения с помощью ростомера и сантиметровой ленты с
* 11

|
|
Рис. 4. Калиперы различной конструкции
точностью до 0,5 см. Калипер обеспечивает точность до 0,5 мм. Вес тела определяется с точностью до 50 г. Все

Рис. 5. Динамометр ручной: а — для взрослых; б — для детей
Рис. б. Динамометр становой:
1 — динамометр; 2 — рукоятка с крюком; 3 — цепь; 4—планка с крюком; 5 — доска для закрепления планки с крюком
инструменты и приборы необходимо периодически про- верять в учреждениях стандартизации метрологии.
Определение антропометрических точек на теле
Для правильного обеспечения антропометрических измерений используют определенные антропометрические точки (рис. 7).
Антропометрические точки на голове
Рис. 7. Основные антропометрические точки тела: v — vertex; г — radiale; tr — trichion; от — omphalion; n — nasion; ic…Антропометрические точки на туловище
Среднегрудинная, mesosternale (mst) — точка в области тела грудины на уровне верхнего края IV грудинно-реберного сочленения (по медиальной… Сосковая, thelion (th) — точка в центре соска; определяется обычно только у… Пупковая, omphalion (от) — точка центра пупка.Антропометрические точки
Плечевая, akromion (a) — наиболее выступающая кнаружи точка на крае акромиального отростка лопатки.Антропометрические точки на черепе
левой глазницы располагались в горизонтальной плоскости. Основные точки следующие (рис. 8):Измерение продольных размеров тела
ность между высотой их стояния и составит искомую величину. Парные точки измеряются на правой стороне. Измерения производят начиная сверху.… Длина тела (рост) — высота над полом верхушечной точки. Обследуемый становится… Длина туловища: верхнегрудинная—лобковая точка.Измерение диаметров
Переднезадний и,поперечный диаметры головы измеряют малым толстотным циркулем. При вычислении переднезаднего диаметра находят расстояние между… Измерения диаметров тела • Акромиальный диаметр (ширинаплеч)-расстояние между правой и левой акромиальными точками..23
• Поперечный диаметр дистальной части предплечья —
наибольшее расстояние между шиловидными отрос
тками лучевой и локтевой костей.
• Поперечный диаметр дистальной части бедра — наи
большее расстояние между медиальным и латераль
ным надмыщелками бедренной кости.
• Поперечный диаметр дистальной части голени — наи
большее расстояние между выступающими точками
лодыжек большеберцовой и малоберцовой костей.
Измерение обхватных размеров тела
• Окружность головы измеряют наложением ленты спереди на уровне глабеллы, сзади — по затылочной точке. • Обхват шеи — лента накладывается горизонтально, спереди она проходит под… • Обхват груди — определяют в 3 состояниях: спокой ного дыхания (пауза), максимального вдоха и макси мального…Определение веса тела
Перед взвешиванием весы проверяют и регулируют. При взвешивании обследуемый становится на середину площадки весов, стоит спокойно. Взвешивание детей… Определить поверхность тела в квадратных метрах без вычислений позволяет… 1,8 1,9 2.0 2,1 22 U 2,4 1J6 Ч ч …ЮО
Определение поверхности тела
S = 71,84 ■w°-435-K0-72S, raeS — площадь тела, см2; Н—длина тела, см; W — мас са тела, кг. - Площадь поверхности тела также можно вычислить поформулеИзмерение кожно-жировых складок (калиперометрия)
Толщина подкожной жировой складки измеряется с помощью калипера или скользящего циркуля (штангенциркуля). Во избежание ошибок тщательно определяют место измерения. Важно правильно поднять кожную складку (рис. 11). Для этого она плотно зажимается большим и указательным пальцами или тремя пальцами так, чтобы в составе складки оказалась бы кожа и подкожный жировой слой. Жалобы на боль свидетельствуют о том, что захвачена только кожа. Пальцы располагают прибли-
To
Таблица 1
Фактор массы тела по В.В. Бунаку
Окончание табл. 1 Р, кг /(Р) Р,кг /(Р) … Таблица 232 . .
Зак. 486
'* 33 3) на передней поверхности плеча в верхней трети внут ренней поверхности… 4) на предплечье на передневнутренней поверхности в наиболее широком его месте, складка берется верти кально; :Глава2. ЧАСТНАЯ
МОРФОЛОГИЯ
Голова
Голова, как и череп, делится на два отдела: мозговой и лицевой. Для характеристики размеров и формы мозгового отдела используют два диаметра: продольный и поперечный. Продольный диаметр — наибольшая длина головы — измеряется от глабеллы до затылочной точки. Его групповые средние варьируют в пределах 172—198 мм. Поперечный диаметр — наибольшая ширина головы — измеряется между теменными точками. Групповые средние варьируют в пределах 134—162 мм. Большое значение в антропологии придают вычислению головного указателя:
поперечный диаметр • 100

продольный диаметр

долихокранный мезокранный брахикранный
Рис. 14. Форма черепов с различным головным указателем
Весь диапазон изменений головного указателя делят на три группы: долихокефалия — ниже 75,9; мезокефалия — 76,0—80,9; брахикефалия — выше 81,0 (рис. 14). Групповые средние головного указателя колеблются в пределах 71—89.
Головной мозг
Индивидуальные и групповые колебания абсолютного веса мозга современных взрослых людей очень велики. Средние групповые значения лежат между 1 100… 1 800 г. Диапазон крайних индивидуальных значений еще шире: от 2 012 г (у И.С. Тургенева) до 1 017 г (у Анатоля…Глазная область
Наружная поверхность век покрыта очень тонкой кожей. В месте перехода ко.жи верхнего века в кожу крыши I? глазницы образуется надглазничная… Кожа верхнего века в одних случаях натянута плотно, не образуя складок, в… Кроме рассмотренных складок верхнего века, различается еще особая складка у внутреннего угла глаза, в боль-; шей или…Область носа
Форма носа обусловлена строением как его костной основы, так и хрящей и мягких тканей. Костный скелет образован лобными отростками верхнечелюстных… Форма носа определяется формой отдельных его элементов: переносья, спинки,… Носовой указатель (процентное отношение ширины к длине при измерении длины от назиона) варьирует по группам от 60 до…Губы
Характерные особенности ротовой полости человека, видимо, возникли в связи с общей перестройкой лицевого отдела головы, в частности с его укорочением и сужением. Эти преобразования, в том числе и формирование губ человека, были тесно связаны с развитием членораздельной речи.
В толще губ находится круговая мышца рта, в которую вплетаются многие мимические мышцы. Снаружи губы покрыты кожей, изнутри — слизистой оболочкой. Между ними находится переходная зона. Она имеет кожный покров, но без рогового слоя. Переходная зона имеет богатую иннервацию и очень чувствительна. От кожи она отличается отсутствием волос и потовых желез, а от слизистой — отсутствием слизистых желез. В ней имеются только мелкие сальные железы. На границе кожи и переходной зоны образуется кайма, иногда принимающая форму валика, например, у негров при вздутых губах. Переходную зону верхней и нижней губ в антропологии принято называть «слизистыми губами». Посредине кожной
части верхней губы сверху вниз опускается бороздка, по бокам ограниченная валиками.
Толстые, выступающие губы с хорошо выраженной переходной зоной и вертикальной бороздкой на верхней губе характерны только для человека. У обезьян, включая антропоморфных, губы тонкие, плотно натянуты на альвеолярных отростках челюстей, повторяют их форму и лишены собственного рельефа; бороздка отсутствует. Переходная зона губ у обезьян не выражена, только у антропоморфных обезьян появляются узкие полоски промежуточных губ. Для антропоморфных обезьян характерна большая высота верхней губы и большая ширина ротовой щели. У человека углы ротовой щели не заходят дальше пред-коренных зубов.
Высокая верхняя губа (19—21 мм) характерна для групп с высоким лицом, например для многих народов Северной Европы, Сибири, Дальнего Востока, Северной Америки, хотя строгой связи между высотой лица и этим признаком не обнаружено. Низкая губа (13—15 мм) чаще встречается у низколицых групп в Южной Европе, на Кавказе, в Передней Азии, Индии, Индокитае, Индонезии, Африке.
Толщина губ — это суммарная высота слизистой части верхней и нижней губ. Тонкими считаются губы до 6 мм, средними — 6—10 мм, толстыми —• 10—13 мм, вздутыми — свыше 13 мм. Нижняя губа обычно толще верхней. У женщин губы тоньше, чем у мужчин. Наибольшая толщина губ у африканцев, австралийцев, меланезийцев, у многих групп Южной Индии, Индокитая, Индонезии, наименьшая у народов Северной Африки и Северной Европы. Толщина губ зависит в основном от степени развития круговой мышцы рта и особенно ее краевых пучков, срастающихся со слизистой. В меньшей степени она зависит от степени жироотложения.
Прохейлия (выступание верхней губы вперед) характерна для африканцев, меланезийцев, австралийцев и мно-
V 41
гих монголоидных групп. Для европеоидов типична орто-хейлия. Выступание верхней губы в известной мере обусловливается прогнатизмом и продентией. Однако у многих монголоидных групп наблюдается прохейлия при отсутствии выступания вперед челюстей и зубов. Описто-хейлия (отступание губы назад) встречается редко, только у отдельных вариантов в европеоидных группах.
Контур верхней губы может иметь разную форму. У негров он вогнутый, у северных европейцев — прямой^ у африканских пигмеев и семангов — выпуклый.
Ушная раковина
ж *& !■-Покровы тела
Кожа
Кожа человека состоит из двух слоев: верхнего слоя — эпидермиса и дермы. Ниже расположен слой подкожной жировой клетчатки. В эпидермисе располагается пигмент кожи — меланин. В толще дермы находятся сальные и потовые железы.
Особый раздел морфологии (дерматоглифика) изучает рельеф кожи.

|
Различают флексорные борозды — линии, приходящиеся на линии сгибов, нтактилъные, нишпапиллярные узоры. Тактильные узоры образованы системой гребешков и бороздок. Гребешки — это папиллярные линии, соответствующие сосочкам дермы — местам, где дерма вдается в эпидермис. Вьщеляют три основных узора папиллярных линий: дуги, петли и завитки, или круги (рис. 17).

Рис. 17. Папиллярные узоры пальцев: 1 — дуга; 2 — петля; 3 — завиток
Волосы
В строении волоса различают корень — часть волоса, расположенную в толще кожи, и стержень — ту часть, которая выступает над кожей. В стержне волоса вьщеляют сердцевину, или мозговое вещество, корковый слой и наружный слой — кутикулу.
'Ч
В течение жизни у человека последовательно появляются три типа волосяного покрова: первичный, или зародышевый, вторичный и третичный.
Первичный волосяной покров, или лануго, выражен
у плода и к моменту рождения сменяется вторичным,
или детским. Третичный волосяной покров появляется к
началу полового созре- /

|
вания на определенных участках тела. Это волосы в области лобка и подмышечной впади- а< ны, а у мужчин на лице (усы и борода), на животе, груди и конечностях. Зародышевые волосы самые тонкие ^-менее 0,03 мм в диа- б< метре. Вторичный волосяной покров образован волосами толщиной 0,03—0,05 мм, которые нркрывают все тело. Волосы головы, брови и ресницы также в< относятся к вторичному волосяному покрову, но имеют большую толщину.
| Рис. 18. Форма волос головы: прямые волосы: а — 1 — тугие; 2 — гладкие; 3 — плосковолнистые; б — волнистые: 4— широковолнистые; 5 — узковолнистые; 6 — локоновые); в — курчавые: 7— завивающиеся; 8 — слабокурчавые; 9 — сильнокурчавые |
Важным признаком является форма волос головы. Различают три основных типа волос: прямые, волнистые и курчавые и несколько . подтипов (рис. 18).
| Большое значение в этнической антропологии придают степени развития бороды (рис. 19). |

Рве. 19. Степениразвития бороды: 1 — очень слабо; 2 — слабо; 3 — средне; 4—сильно; 5—очень сильно
Пигментация
В коже меланин образуется в клетках росткового слоя эпидермиса. Меланин может быть распределен диффузно или в виде зерен. Диффузный меланин сдвигает… В волосах пигмент сосредоточен в корковом слое. Отечественные антропологи… Цвет глаз определяется количеством и глубиной залегания пигмента в радужке. Если пигмент сосредоточен в глубоких…Зубы
У человека, как и у большинства млекопитающих, дифидонтная зубная система — последовательно прорезываются два поколения зубов: на смену 20 молочным зубам приходят 32 постоянных зуба (рис. 20). Зубная формула для постоянных зубов у человека:
3212:2123
3212:2123 '
Или I
{dentes canini) — клыки; Р (dentes premolares) — малые коренные зубы; М (denies molares) — большие коренные зубы. Рп — премоляр нижней челюсти; Мп— моляр верхней челюсти.
Общепринятой теорией происхождения зубов у мле-хопитающих, в том числе зубов приматов, является три-
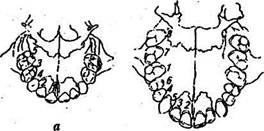
Рис.20. Порядок прорезывания зубов: а — молочных, б — коренных
* 47
А: -
Ял
| рлгвсяят |
| tntwniu |
| metacmiu |

 туберкулярная теория, основанная Коном и развитая Ос-борном. Коронка простого зуба рептилий приобретала более сложное строение благодаря появлению на ее верхушке, наряду с основным конусом — проконусом — еще двух добавочных: переднего, или параконуса, и заднего, или метаконуса (для нижнего зуба названия кончаются на «ид»: протоконид, параконид и метаконид), получилась стадия трикодонтных зубов.
туберкулярная теория, основанная Коном и развитая Ос-борном. Коронка простого зуба рептилий приобретала более сложное строение благодаря появлению на ее верхушке, наряду с основным конусом — проконусом — еще двух добавочных: переднего, или параконуса, и заднего, или метаконуса (для нижнего зуба названия кончаются на «ид»: протоконид, параконид и метаконид), получилась стадия трикодонтных зубов.
В дальнейшем подобные зубы усложнились в своем строении в связи с общим ходом эволюции их обладателей и их способом питания и приобрели весьма разнообразные формы в разных группах млекопитающих. При усложнении коронки число корней увеличивалось в некотором, но не в полном, соответствии с размером усложнения верхней части зуба.
По мере разрастания и повышения пятки на ней появляются новые бугорки, и получаются пяти- и шестибу-горчатые зубы: на тритоне между протоконусом и парако-нусом вдвигается добавочный бугорок — протоконулюс, а между протоконусом и метаконусом — добавочный бугорок — метаконулюс; на талониде позади метаконида формируется основной бугорок — энтоконид, между последним и гопоконидом — добавочный бугорок гипоко-нулид, иначе, по своему положению посредине заднего отдела талонида у современного человека и у некоторых других приматов, называемый мезоконидом. Наконец, у некоторых дриопитеков, у оранга и современных людей между энтоконидом и гипоконулидом встречается еще один добавочный бугорок — «шестой бугорок».
Приведем список названий бугорков с их общепринятыми сокращенными обозначениями согласно тритубер-кулярной теории Кона—Осборна (рис. 21).
Бугорки наверхних молярах:
Передний наружный — параконус (paraconus — pa).
Передний промежуточный — протоконулюс (proto-conulus — pi) у человека отсутствует.
| Ъдросвпю |
| prfftocgnus |
ЬуросопиШ pntoconid
Рис. 21. Бугорки на верхних и нижних молярах человека
Передний внутренний — протоконус (ргрюсопш — рг).
Задний наружный — метаконус (metaconus — me).
Задний промежуточный — метаконулюс (metaco-nulus — ml) у человека отсутствует.
Задний внутренний —гипоконус (hypoconus — hy).
Бугорки на нижних молярах:
Передний внутренний — параконид (paraconid — pad) учеловека отсутствует.
Передний наружный — протоконид (protoconid—prd).
Задний внутренний — метаконид (metaconid — med).
Задний наружный — гипоконид (hypoconid — hyd).
Самый задний внутренний — энтоконид (entoconid — end). .
Задний промежуточный средний— гипоконулид (hyppconulid — hid) или мезоконид (mesoconid — msd).
Задний добавочный внутренний — шестой бугорок (tuberculum sextum — 6) или бугорок Кирабелли встреча- ется не всегда.
 | |||
 | |||
Глава3. ВОЗРАСТНАЯ
АНТРОПОЛОГИЯ
 Возрастная антропология изучает закономерности становления и развития анатомических структур и физиологических функций на протяжении онтогенеза — от оплодотворения яйцеклетки до конца жизни.
Возрастная антропология изучает закономерности становления и развития анатомических структур и физиологических функций на протяжении онтогенеза — от оплодотворения яйцеклетки до конца жизни.
Рост и развитие организма
Существуют два вида морфологических исследований процесса роста у человека: продольные и поперечные. При продольных исследованиях… явить взаимосвязь морфологических и функциональных показателей, а также понять… Процесс роста и развития организма характеризуется следующими закономерностями:Общая периодизация онтогенеза
Детальная схема периодизации онтогенеза человека была предложена В.В. Бунаком в 1965 г. (табл. 3). По этой схеме весь период онтогенеза делится на… Сходная схема (табл. 4) возрастной периодизации пост-натального развития… Пренатальный период подразделяется на два: эмбриональный и фетальный (плодный). В течение первого пери-Таблица 3
Схема периодизации индивидуального развития (В.В. Бунак, 1965)
| Стадия | Период | Возраст | Пол |
| мужской | женский | |||
| Внутриутробный цикл | |||
| эмбриональный переходный фетальный | ранний средний поздний | 0-8 нед. 8-16 нед. 4-6 мес. 7-8 мес. 8-10 мес. | |
| Внеутробный цикл | |||
| начальный | 1-6 мес. | ||
| Прогрессивная | младенческий | средний конечный | 7-9 мес. 10-12 мес. |
| первого детства | начальный конечный | 1-4 года 5-7 лет | |
| второго детства | начальный конечный | 8-10 лет 8-9 лет 11—13 лет 10-12 лет | |
| подростковый | 14-17 лёт 12-16 лет | ||
| юношеский | 18-21 лет 17-20 лет | ||
| Стабильная | взрослый | первый второй | 22-28 лет 21-26 лет 29-35 лет 27-32 года |
| зрелый | первый второй | 36-45 лет 33-40 лет 46-55 лет 41-50 лет | |
| пожилой | первый второй | 56-63 года 51-57 лет 64-70 лет 58-63 года | |
| Регрессивная | старческий | первый второй | 71-77 лет 64-70 лет 78-83 года 70-77 лет |
| позднестарческий | 84 года 78 лет |
ода, который продолжается 8 недель, происходит форми- рование органов и частей тела, свойственных взрослому че-- ловеку. В фетальный период главным образом увелйчива-i размеры и завершается органообразование. Скорость i плода возрастает до 4—5 месяцев. После 6 месяцев
53
Таблица 4 Схема возрастной периодизации онтогенеза человека
| 1. | Новорожденные | 1-10 дней |
| 2. | Грудной возраст | 10 дней — 1 год |
| 3. | Раннее детство | 1-3 года |
| 4. | Первое детство | 4-7 лет |
| 5. | Второе детство | 8-12 (мальчики) 8-11 (девочки) . |
| 6. | Подростковый возраст | 13—16 (мальчики) 12-15 (девочки) |
| 7. | Юношеский возраст | 17—21 (юноши) 16-20 (девушки) |
| 8. | Зрелый возраст I период | 22-35 (мужчины) 21-35 (женщины) |
| II период | 36-60 (мужчины) 36-55 (женщины) | |
| 9. | Пожилой возраст | 61-74 (мужчины) 56-74 (женщины) |
| 10. | Старческий возраст | 75-90 лет (мужчины и женщины) |
| П. | Долгожители | 90 лет и выше |
скорость роста линейных размеров уменьшается. По-видимому, одна из причин замедления роста в конце внутриутробного периода — ограниченные размеры полости матки. По данным Дж. Таннера, скорость роста близнецов замедляется в тот же период, когда их общий вес становится равным весу одиночного 36-недельного плода.
Сразу после рождения наступает период, называемый периодом новорожденности. Основанием для его выделения служит тот факт, что в это время имеет место выкармливание ребенка молозивом в течение 8—10 дней.
Следующий период — грудной — продолжается до 1 года. Начало его связано с переходом к питанию «зрелым» молоком. Во время грудного периода наблюдается наибольшая интенсивность роста по сравнению со всеми
остальными периодами внеутробной жизни. Длина тела увеличивается от рождения до года примерно в 1,5 раза, а вес утраивается. С 6 месяцев начинают прорезываться молочные зубы.
Период раннего детства длится от 1 года до 3 лет. На 2—3-м году жизни заканчивается прорезывание молочных зубов. После 2 лет абсолютные и относительные величины годичных приростов размеров тела быстро уменьшаются. . С 4 лет начинается период первого детства, который 'заканчивается в 7 лет. В этот период некоторые исследователи отмечают небольшое увеличение скорости роста, на-зывая его первым ростовым скачком; однако было обращено внимание, что этот скачок свойственен не всем детям.
I Начиная с 6 лет появляются первые постоянные зубы: пер-
ШЧиый моляр и центральный резец на нижней и верхней че-к., латеральный резец на нижней челюсти.
>{ ,„, Возраст от 1 года до 7 лет называют также нейтраль-;вым детством, поскольку мальчики и девочки почти не
* "отличаются друг от друга по размерам и форме тела. Сле-%Дует отметить, что уже в этот период количество жира у девочек больше.
| J |
Период второго детства длится у мальчиков с 8 до 12 лет, у девочек — с 8 до 11 лет. В этот период выявляются половые различия в размерах и форме тела, а также ^йачинается усиленный рост в длину. Темпы роста у девочек выше, чем у мальчиков, так как половое созревание у $евочек начинается в среднем на два года раньше. Примерно в 10 лет девочки обгоняют мальчиков по длине и весу тела, ширине плеч. В этот период у девочек быстрее ^растут нижние конечности, происходит интенсивное увеличение показателей массивности скелета. В среднем к 12—13 годам у мальчиков и девочек заканчивается смена зубов.
В период второго детства повышается секреция полого "- ^«гормонов, врезультатечегоначинаютразвиватьсявто-' f- ричные половые признаки. Последовательность появле-
'*- 55
ния вторичных половых признаков довольно постоянна: у
девочек сначала формируется грудная железа, затем по
являются волосы на лобке, а потом в подмышечных впа
динах. Матка и влагалище развиваются одновременно с
формированием грудных желез. Средний возраст разви
тия грудных желез у девочек различных этнических групп
колеблется от 9 до 10 лет. Средний возраст появления во
лос на лобке приходится на самый конец периода второго
детства. к
В гораздо меньшей степени в этот период процесс полового созревания выражен у мальчиков. Лишь к концу периода второго детства у них начинается ускоренный рост яичек, мошонки, а затем полового члена. По данным болгарских медиков, длина полового члена в возрасте от 8 до 10 лет практически не меняется и увеличивается к 12 годам всего на 0,7 см.
Следующий период — подростковый — называют также периодом полового созревания, или пубертатным. Он продолжается у мальчиков с 13 до 16 лет, у девочек^— с 12 до 15 лет. Датировку этого периода нельзя считать окончательной, поскольку по уровню полового созревания 13-летние мальчики соответствуют не 12-, а 11-летним девочкам. Поэтому у мальчиков к началу подросткового периода только начинается половое созревание, напротив, у девочек оно в значительной степени захватывает еще и предшествующий период. В этот период наблюдается дальнейшее увеличение скоростей роста — пубертатный скачок, который касается всех размеров тела. Наибольшие прибавки по длине тела у девочек имеют место между 11 и 12 годами, по весу тела — между 12 и 13 годами; у мальчикоасоответ-ственно между 13 и 14 и 14 и 15 годами. Особенно велики скорости роста большинства размеров у мальчиков, в результате чего к 13,5—14 годам они обгоняют девочек по длине тела. К концу подросткового периода размеры тела составляют 90—97% своей окончательной величины.
В подростковый период происходит перестройка основных физиологических систем организма (мышечной, кровеносной, дыхательной и др.). К концу периода основные функциональные характеристики подростков приближаются к характеристикам взрослого организма. У мальчиков в это время особенно интенсивно развивается мышечная система.
В подростковый период формируются вторичные половые признаки. У девочек продолжается развитие грудных желез, рост волос на лобке и в подмышечных впадинах. Наиболее четким показателем полового созревания женского организма является менструация (менархе). Она обычно начинается после того,' как пройден максимум скорости роста тотальных размеров тела. В 70—80-х гг. возраст появления менархе у городского населения большинства европейских стран был около 13 лет. У девочек, живущих в сельской местности, отмечены более поздние сроки менархе: разница с городским населением составляет от 6 до 10 месяцев. Наиболее поздние сроки первой менструации характерны для подростков, живущих в экстремальных условиях обитания. Например, у девочек высокогорья Киргизии первая менструация наступает в 15 лет, что связывают с адаптацией к гипоксии.
В подростковый период происходит интенсивное половое созревание у мальчиков. Продолжается рост яичек и полового члена, особенно интенсивно в 13—14 лет. К 13 годам происходит мутация голоса и появляются волосы на лобке; к 14 годам наблюдается пубертатное набухание сосков и появляются волосы в подмышечных впадинах. К 15 годам начинается рост волос на верхней губе и подбородке. В14—15 лет у мальчиков появляются первые поллюции (непроизвольное извержение семени).
Следует заметить, что у мальчиков по сравнению с девочками более продолжителен предпубертатный период и сильнее выражен пубертатный скачок. Дж. Таннер считает, что «различия в размерах тела между взрослыми
мужчинами и женщинами в значительной степени зависят от времени наступления, продолжительности и интенсивности пубертатного скачка роста. До этого скачка различия в росте между мальчиками и девочками не превышают 2%, а после него они достигают в среднем 8 %».
Юношеский возраст продолжается у юношей от 18 до 21 года, у девушек — от 17 до 20 лет. В этот период в основном заканчивается процесс роста и формирования организма, и все основные размерные признаки достигают дефинитивной (окончательной) величины.
В зрелом возрасте форма и строение тела изменяются мало. Правда, у 20—30-летних людей еще продолжается рост позвоночного столба за счет отложения новых слоев костного вещества на верхних и нижних поверхностях позвонков. Однако этот рост незначителен и не превышает в среднем 3—5 мм. Между 30 н 45—50 годами длина тела остается постоянной, а потом начинает уменьшаться. В пожилом и старческом возрасте происходят инволютивные изменения организма.
Биологический возраст
Формулирование понятия биологический возраст имеет большое значение, поскольку для многих практичес- ких целей важна группировка детей не по календарному (паспортному) возрасту, а… Основнымикритериями биологического возраста счи-. таются: 1) зрелость, оцениваемая постепени развития вторичных…Вторичные половые признаки
' Ни развития вторичных половых признаков, поскольку это наиболее доступная оценка при массовых обследованиях.Скелетный возраст
ным из всех критериев био- 2..................... логического возраста явля- елся скелетный возраст, по- 5 ...................................... 8Зак. 486
У новорожденных мальчиков и девочек масса мозга составляет в среднем 353 и 347 г соответственно, она примерно удваивается к 6 месяцам (600 и 534 г)… Проблема взаимоотношений биологического возраста с эмоциональным и умственным… детельствуют ассоциации между степенью миелинизации и психоэмоциональным статусом детей. Выделение зависящих от…Видовая продолжительность жизни
Не существует однозначного определения видовой продолжительности жизни. Условно под ней понимается тот возраст, до которого потенциально могут… ■*. ■ 71Глава 4. КОНСТИТУЦИОНАЛЬНАЯ АНТРОПОЛОГИЯ
 Конституция человека — совокупность функциональных и морфологических особенностей организма, сложившихся на основе наследственных и приобретенных свойств и определяющих реактивность организма на различные воздействия. Строение и функциональные особенности организма у различных людей в какой-то мере могут быть сходными, что позволяет говорить о типах конституции человека. Чаще конституция человека определяется по телосложению — совокупности внешних признаков: рост, вес, степень развития мускулатуры и подкожного жирового слоя, пропорциональность отдельных размеров тела, которые устанавливаются антропометрическими измерениями.
Конституция человека — совокупность функциональных и морфологических особенностей организма, сложившихся на основе наследственных и приобретенных свойств и определяющих реактивность организма на различные воздействия. Строение и функциональные особенности организма у различных людей в какой-то мере могут быть сходными, что позволяет говорить о типах конституции человека. Чаще конституция человека определяется по телосложению — совокупности внешних признаков: рост, вес, степень развития мускулатуры и подкожного жирового слоя, пропорциональность отдельных размеров тела, которые устанавливаются антропометрическими измерениями.
Пропорции тела
'А Пропорции тела определяются в первую очередь размерами скелета. Однако на… Наиболее распространенным и доступным широкой практике способом оценки пропорций тела человека является метод…Таблица б Характеристика пропорций тела (по Башкирову, 1937)
| Типы пропорций тела | Размеры тела, в процентах от длины тела | ||||
| Длина туловища | Длина ноги | Длина руки | Ширина плеч | Ширина таза | |
| Долихоморфный Мезоморфный Брахиморфный | 29,5 31,0 33,5 | 55,0 53,0 51,0 | 46,5 44,5 42,5 | 21,5 23,0 24,5 | 16,0 16,5 П,5 |


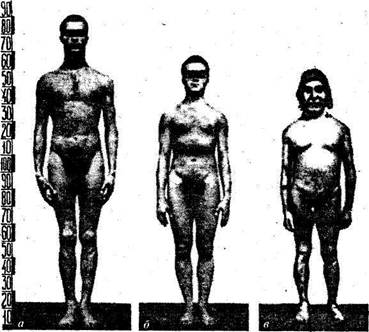
Рис. 24. Типы конституции человека: а — долихоморфный; б — мезоморфный; в — брахиморфный
Пропорции тела, выявленные с помощью индексов, могут сравниваться лишь при сходной длине тела. Если же между сравниваемыми индивидами (или группами) различия по общей длине тела значительны, то метод индексов не позволяет судить с достаточной точностью о существующих различиях в пропорциях тела, так как природа индекса не учитывает зависимости входящих в него размеров тела от его общей величины. Известно, что с увеличением длины тела человека длина ноги увеличивается относительно быстрее, а продольные и поперечные размеры туловища медленнее. В результате люди более высокорослые, независимо от принадлежности к тому или иному типу пропорций тела, окажутся относительно более длинноно-
гими, с более коротким и узким туловищем, и наоборот. Таким образом, для точной сравнительной характеристики пропорций тела необходимо учитывать существующиемор-фогенетические корреляции роста различных его частей.
Используя корреляционный и регрессивный анализ размеров тела и, опираясь на невысокую корреляцию поперечных размеров туловища с длиной конечностей, В.В. Бунак разработал типологию пропорций тела, состоящую из 9 основных вариантов:
1) арростоидный (arrostos — слабосильный) — коротко
ногий и узкоплечий;
2) гармоноидный — средний по длине ноги и ширине
плеч;
3) гигантоидный — длинноногий и широкоплечий;
4) гипогармоноидный — узкоплечий при средней дли
не ног;
5) парагармоноидный — широкоплечий при средней
длине ног;
6) гипостифроидный (stiphros — крепкий, плотный) —
коротконогий при средней ширине плеч;
7) стифроидный — коротконогий и широкоплечий;
8) тейиоидный (teino — вытянутый) — длинноногий и
узкоплечий;
9). паратейноидный — длинноногий со средней шириной плеч. Учет изменчивости других размеров — длины руки
ширины таза, сегментов конечностей — дает возможность
выделить ряд подтипов.
Возрастная изменчивость пропорций тепа
степенно видоизменяются за счет разной скорости роста отдельных частей (рис. 25). . * В допубертатном периоде развития дети характеризуются относительно короткими… В связи с разной интенсивностью роста размеров тела в пубертатном периоде подростки по сравнению с детьми.Половые различия в пропорциях тела
Рис. 26. Пропорции мужского и женского тела (по Грицеску) v 89Состав тела
-; В прошлом анатомы расчленяли трупы и определяли процентное содержание веса тканей и отдельных органов по отношению к общему весу тела. Знание… Анализ вариаций клеточной массы у европеоидов в возрастном интервале от 8 до…Морфологические аспекты конституции
в^ялую и упругую и рекомендовал принимать во внимание конституционные особенности при лечении болезней. Позднее Гален ввел понятие о габитусе, т.е.… Среди специалистов нет единого мнения о человеческой конституции и условно… Некоторые специалисты предлагают различать общую и частные конституции. Общая конституция есть общее суммарное…Й
"4*
Рйс. 30. Типология Сиго:
а — мускульный; б—церебральный;» — респираторный; г—деигестивный
 | |||||
 | |||||
 |

|

Рис. 31. Типология Кречмера
бедра. Пропорции и характерные особенности диспласти-ка трудно поддаются описанию.
Женские конституции
А. Лептосомные конституции 1. Астенический тип характеризуется худым телом, с плоской, узкой, длинной грудной клеткой, втянутым животом, узким тазом, с длинными ногами; междуДетские конституции
Так, в России широко применяется схема В.Г. Штеф-ко и А.Д. Островского (1929), пригодная для классификации и взрослых. В ней описаны нормальные… Среди нормальных типов выделяются: 1. Торакальный — с сильным развитием грудной клет ки в длину, небольшим животом, большой жизнен ной емкостью легких…Психофизиологические аспекты конституции
строением тела, физиологией жизненных функций и метаболизмом, представляющими лабильный и динамичный феномен, и разумом, сознанием —… 4.4.1. Конституции и физиологические особенности . В мировой литературе крайне мало данных по физиологическим особенностям отдельных типов конституции. Считается, что у…Подотряд полуобезьяны (Prosimii)
риновых приматов, которые обладают ноздрями в виде запятых, открывающимися на оголенную часть кончика носа. Верхняя губа у этих приматов гладкая… Подотряд полуобезьян объединяет 6 семейств, 26 родов и около 50 видов с…Семейство тупайеобразные (Tupaiidae)
Зубная формула: 3312:2133 3313:3133V из
 Питаютсятупайи преимущественно в»^комыми. Ту-пайи широко распространены в тропических дождевых и горных лесах (свыше 3000м над уровнем моря) Северной Индии, Южных Гималаев, Китая, Индокитая, на островах Хайнань, Суматра, Калимантан и Бали.
Питаютсятупайи преимущественно в»^комыми. Ту-пайи широко распространены в тропических дождевых и горных лесах (свыше 3000м над уровнем моря) Северной Индии, Южных Гималаев, Китая, Индокитая, на островах Хайнань, Суматра, Калимантан и Бали.
Семейство Tupaiidae подразделяется на два подсемейства: 1) собственно тупайи (Tupaiinae) с родами Tupaia, Dendrogale, Urogale и Anathana и 2) перохвостые тупайн (Pttticercinae) е одним родом Ptilocercus. В отличие от других, тупаий перохвостые тупайи ведут ночной или сумеречный образ жизни.
5.1.2. Семейство лемуры (Lemuridae)
Семейство лемуряд, или лемурообразных полуобе-зьян, объединяет собственно лемуров, обитающих на Мадагаскаре и некоторых мелких соседних с ним островах.
У этих животных Густой волосяной покров е разнообразной раскраской, хвост длинный» пушистый; мордочка чаще удлиненная, как у лисицы; имеется 4—5 групп осязательных волос —вибрисс, глаза крупные, сближенные. Конечности хватательные с хорошо противопоставленными большими пальцами. На всех пальцах — ногти, только на втором пальце стопы коготь, который называется «туалетным»: он служит для расчесывания шерсти.
Зубная формула:
33 12:2133
Преимущественно лемуры растительноядные, но некоторые виды питаются также насекомыми и яйцами птиц.
Семейство Lemuridae разделяется на два подсемейства: лемуровьге, или истинньге лемуры (Lenmrinae) с родами Lemur, Hapalenrar, Lepdlemur и мышиные лемуры (Cheirogateinae) с родами Cherogalens, Microcebus, Phaher.
5.1.3. Семейство индризиды {indrisidae)
Индризиды имеют длинные задние конечности, с помощью которых передвигаются по земле большими прыжками, при этом передние лапы вытягивают вверх или вперед. Пальцы с тыльной стороны покрыты волосами; на ногах они соединены кожной перепонкой до вторых фаланг, но большой палец свободный и противопоставляется остальным, на кистях первый палец маленький, который слабо противопоставляется. Все пальцы имеют i ногти, а на втором пальнеяога—коготь. Зубная формула:
3212:2123
 3 202:2023
3 202:2023
ной пищей. Известны виды как с дневным, так и ночным |f образом жизни.
К этому семейству относятся три рода: черных ко-| роткохвость1хиядри, илибабакото(Ьк}г1), мохнатых инд-
ри, или авагисов (Avahi), и хохлатых индри, или сивак |, (Propithecus).
 33 12:2133
33 12:2133
На верхней челюсти срединные резцы широко расставлены (диастема), нижние резцы вместе с клыками сближены и сильно наклонены вперед, образуя «зубной гребешок». Имеется нижний язычок. Лемуриды ведут дневной, ночной и сумеречный образ жизни. Есть древесные, полудревесные и наземные формы. Слово «лемур» означает «привидение», «дух усопшего».
5.1.4. Семейство руконожковые (Daubenioniidae)
Руконожковых называют также шл-ойл. Этих животных в 1780 г, открыл путешественник Пьер Соняера на £• западном берегу острова Мадагаскар. Сами мадагаскар-I ты, которым Соннера показал пойманных зверьков, нн-> «огда не видели их раньше и громко кричали от удивле-
^ 115
 ния, и Соннера эти возгласы «айл-айл» избрал в качестве названия для открытого им существа.
ния, и Соннера эти возгласы «айл-айл» избрал в качестве названия для открытого им существа.
К семейству руконожковых относится только род Daubentonia с одним видом — руконожка мадагаскарская (D. madagascariensis). Руконожка размером с кошку: длина туловища и головы около 40 см, а хвост длиннее (около 60 см). Конечности короткие, причем задние длиннее передних. На всех пальцах — когти, только на крупном большом пальце ноги имеется настоящий плоский ноготь. На передних конечностях средний палец очень тонкий и длинный.
Благодаря особенностям в зубной системе руконожек когда-то причисляли даже к грызунам. Крупные передние зубы верхней и нижней челюстей айл-айл постоянно растут и покрыты эмалью только на передней стороне. Зубная формула:
3101:1013
 3 001: 100 3
3 001: 100 3
Айл-айл живут в бамбуковых зарослях и на крупных ветвях и стволах деревьев дождевых лесов Мадагаскара. Встречаются одиночками, реже парами. Питаются фруктами, включая плоды мангового дерева и кокосовых пальм, сердцевиной бамбука и сахарного тростника, любят также древесных жуков и личинок. Руконожки — типичные ночные животные.
Семейство лоризиды (Lorisidae)
ных родов тонкие и толстые лори обитают в Азии, а перо-диктикусы, арктоцебусы и галаго — в Африке. Лоризиды — древесные и ночные животные. Зубная формула: • 33 12:2133Семейство долгопяты (Tarsiidae)
Долгопяты — маленькие животные. У них большая круглая голова, мордочка широкая и укороченная с очень крупными глазами, смотрящими прямо вперед, как… V 117 3312:2133 3311:1133 *Подотряд человекоподобные высшие приматы (Anthropoidea)
У всех обезьян на пальцах плоские ногти <у игрунко-вых — когтеобразные ногти); глаза обращены вперед, и глазница полностью отделена от височной… Для обезьян характерна редукция обонятельного аппарата и специальных… ко у эдиновых игрунок и в большей степени у ночных обезьян на ладонях и подошвах еще встречаются участки кожи без…Семейство игрункообразных (Callitrichidae)
Иногда игрунок называют белкообразными обезьяна-^^^ ми. Все пальцы игрунок несут сильно сжатые с боков, заостренные и сводчатые ногти, похожие на… 2 3 12: 2 1 3 2 2312:2132V 121


 Все пальцы цепкохвостых обезьян имеют сводчатые ногти. Большой палец стопы хорошо развит и может сильно отодвигаться в сторону. Напротив, большой палец кисти короткий, у некоторых видов сильно редуцирован и даже отсутствует (коата). .Ходят цебиды на четырех конечностях, полностью опираясь на ладонную и подошвенную поверхность.
Все пальцы цепкохвостых обезьян имеют сводчатые ногти. Большой палец стопы хорошо развит и может сильно отодвигаться в сторону. Напротив, большой палец кисти короткий, у некоторых видов сильно редуцирован и даже отсутствует (коата). .Ходят цебиды на четырех конечностях, полностью опираясь на ладонную и подошвенную поверхность.
Волосяной покров густой, окраска его сильно варьирует. Зубная формула:
33 12:2133
 3312:2133
3312:2133
Имеется одна пара сосков. Рождается один детеныш. Горловые мешки и седалищные мозоли отсутствуют. Обитают цебиды в Центральной и Южной Америке. Ведут чисто древесный, дневной, стадный образ жизни.
Семейство цебида подразделяется на подсемейства: ночные обезьяны (Aotinae) с родами Aotus и Callicebus; чертовы обезьяны, или саки (Pithecinae), с родами Pithecia, Chiropotes и Cacajao; ревуновые обезьяны (Alonattinae) с одним родом Alouatta; капуциновые обезьяны (Cebinae) с родами Cebus и Saimiri; паукообразные обезьяны (Atelinae) с родами Ateles, Brachyteles и Lagomrix. Всего в семействе 11 родов, включающих 20 видов.
5.2.3. Семейство мартышкообразные (Cercopithecidae)
В подсемейство мартышковых обезьян входит 8 родов.
Род обыкновенных макак (Масаса) включает животных средних и крупных размеров с массой тела 3,3—18 кг, самки значительно меньше. Для всех представителей этого рода характерно плотное телосложение; конечности почти равной длины; кисть хватательная, большой палец кисти, хотя и короткий, полностью противопоставлен остальным. Между пальцами кистей и стон имеются неболь-
шие кожные перепонки. Шерсть плотная, по окраске варьирует у разных видов. Морда умеренно удлиненная и округленная. Лицо оголенное. Над носом и глазницами имеется толстый костный валик, более выраженный у самцов. Ноздри расположены не на конце морды, а в виде узких носовых щелей, обращенных вперед и вверх. Уши голые, с заостренной верхушкой. Под щитовидной железой расположен непарный горловой мешок, играющий роль резонатора. Защечные мешки хорошо заметны, когда набиты пищей-Длина хвоста у разных видов варьирует.
К этому роду относится 12 видов (46 подвидов), которые широко представлены на севере Африки и в Азии от Пакистана до Японии и Тайваня, от Тибета до Цейлона, а также на островах Малайского архипелага до Филиппин* Встречаются в тропических дождевых и горных лесах (до 4000 м над уровнем моря), в умеренных лесах Китая и Японии, а также в травянистых и сухих областях среди кустарников и кактусов в Индии и на Цейлоне.
Макаки —полудревесные, полуназемные животные. Чтобы уберечься от хищников, они спят на деревьях или прячутся в утесах и скалах. Пища их разнообразна — плоды, корни, молодые листья, насекомые, личинки, моллюски, ракообразные. Они часто совершают набеги на плантации и посевы, где поедают рис, сахарный тростник, кукурузу, картофель.
Поведение и взаимоотношения макак хорошо изучены в природе и в лабораторных условиях. Макаки живут большими группами, в которые входят более чем один самец и несколько самок с детенышами разных поколений. В таких группах ярко выражена иерархия. Центральное место занимает сильный самец, затем самки и детеныши. Иногда подчиненные самцы живут изолированно. 1 Стадо занимает определенную территорию в несколько квадратных километров. Часто ареалы обитания отдельных стад перекрывают друг друга, но драки между стадами бывают редко.
i-. 123


 В соответствии с их сложной социальной жизнью макаки имеют широкий размах эмоций для общения друг с другом. У них отмечено свыше 30 различных звуков. Кроме того, хорошо описано выражение лица — шевеление бровей, улюлюканье губами и т.д.
В соответствии с их сложной социальной жизнью макаки имеют широкий размах эмоций для общения друг с другом. У них отмечено свыше 30 различных звуков. Кроме того, хорошо описано выражение лица — шевеление бровей, улюлюканье губами и т.д.
К роду черных сулавесских хохлатых павианов (Cynopithecus) относится только один вид С. niger с двумя подвидами. Сулавесских павианов иногда считают промежуточной формой между макаками и павианами. Хохлатые черные павианы обитают на острове Сулавеси. Их тело покрыто черной или темно-коричневой густой шерстью. На темени волосы очень длинные, жесткие, торчат вверх в виде хохолка, который у самца развит сильнее. Надглазничный валик очень большой. Лицо черное и оголенное, с обеих сторон носа идут длинные костные гребешки, более сильно развитые у старых самцов. Верхняя губа длинная и широкая. Уши крупные и остроконечные. Седалищные мозоли состоят из двух гладких безволосых розовых подушек. Защечные мешки большие. Конечности почти равной длины, хвост редуцирован до 1—2 см, масса тела около 10 кг.
Род мангобеев, или черномазых обезьян (Cercocebus), распространен в Центральной и Западной Африке. Это древесные, довольно крупные обезьяны, со стройным телом, длинными конечностями и длинным хвостом. Между пальцами имеются кожные перепонки. Череп и морда удлиненные. Седалищные мозоли сливаются в полулунное образование.
Живут мангобеи группами в 20—40 особей, но спят
ночью по 4—5 обезьян вместе. Старые самцы часто живут
в одиночку. Мангобеев считают молчаливыми животны
ми, но у них хорошо развита мимика — оскаливание зу
бов, шевеление губ, бровей, различные угрожающие дви
жения. Питаются мангобеи орехами, семенами, сочными
плодами, листьями. ,
Род павианов (Papia) объединяет 5 видов с 11 подвидами, занимающими широкий ареал по всей Африке, к югу от Сахары, а также в западной части полуострова Аравия.
Павианы — крупные животные, с массой тела до 25 кг и более. Самки почти в 2 раза легче самцов. У павианов длинная морда, что оправдывает их название — собакого-ловые. Клыки сильно развиты. Надбровные валики большие, из-за них глаза смотрят вниз. Ноздри широко открыты и находятся на конце морды.
Живут стадами до 80—100 особей. Такие сообщества павианов характеризуются ярко выраженной иерархией. Каждая группа состоит из взрослых особей, подростков и детенышей. Отношения между членами стада основываются на господстве и подчинении. Господствующее положение принадлежит сильному взрослому самцу.
Стадо павианов занимает обычно 2-^-3 км2. Спят они в расщелинах и под навесами скал, чаще всего сидя, прижавшись друг к другу, детеныши — под защитой матерей. Просыпаются на восходе солнца и начинают активную жизнь в поисках пищи и развлечений.
Павианы всеядны: питаются сочными плодами, молодыми л обегами растений, кореньями, ящерицами, насекомыми, яйцами птиц; совершают набеги на поля, сады и огороды.
К роду мандрилов (Mandrillus) относятся два вида: мандрил (М. sphinx) и дрил (М. leucophaeus). В строении зубов и некоторых других признаков мандрил и дрил сходны с павианами, но во многом и отличаются от них. В.частности, пальцы кистей и стоп более длинные, чем у павианов. Длина хвоста лишь 7 см у мандрил и 12 см у дрил.
Мандрил — одно из наиболее удивительных ярко окрашенных животных среди млекопитающих. Тело мандрила крупное, размером с собаку колли, голова огромная с массивной выступающей мордой. По бокам длинного
V 125



 носа тянутся вздутия, изрезанные продольными бороздами (у дрила они меньше). Цвет вздутий ярко-голубой, а нос, выступающий длинной полоской между ними, и ноздри красные. Щечные хохолки и усы беловатые, борода желтая или оранжевая. Глаза сидят глубоко, уши мясного цвета. Шерсть на теле длинная, густая, на верхней стороне туловища она оливково-коричневатая, на нижней — серебристо-серая. Околохвостовая голая кожа красновато-фиолетовая, седалищные мозоли фиолетовые. Самки мандрилов отличаются от самцов: ©ни в 2—3 раза меяьчесам-цов, в целом менее ярко окрашены, у них отсутствуют синие щечные вздутия.
носа тянутся вздутия, изрезанные продольными бороздами (у дрила они меньше). Цвет вздутий ярко-голубой, а нос, выступающий длинной полоской между ними, и ноздри красные. Щечные хохолки и усы беловатые, борода желтая или оранжевая. Глаза сидят глубоко, уши мясного цвета. Шерсть на теле длинная, густая, на верхней стороне туловища она оливково-коричневатая, на нижней — серебристо-серая. Околохвостовая голая кожа красновато-фиолетовая, седалищные мозоли фиолетовые. Самки мандрилов отличаются от самцов: ©ни в 2—3 раза меяьчесам-цов, в целом менее ярко окрашены, у них отсутствуют синие щечные вздутия.
Обитают мандрилы в Западной Африке. Как и павианы, мандрилы всеядны. Встречаются семейными группами. Мандрилы—агрессивные и драчливые животные.'
Род гелад (Theropithecus) известен из Эфиопии. Гела-ды — крупные обезьяны, массой 20 кг и более; хвост непушистый, с кисточкой волос на кончике; длина хвоста (45—50 см) меньше, чем длина туловища и головы (TOTS см). Шерсть длинная, шелковистая, буроватая. У самцов щечные пучки волос сильно развиты и направлены вниз-назад. Взрослый самец имеет длинную мантию, покрывающую переднюю часть тела. Самки значительно мельче самцов, и мантия у них отсутствует. Интересной особенностью гелад являются бледные голью участки кожи на шее и груди, у самок эти голые участки окружены мясистыми наростами в виде «ожерелья». При возбуждении и в период половых циклов наросты переполняются кровью и становятся багровыми. Кроме того, на груди у самок два соска находятся настолько близко друг к другу, что детеньнн захватывает ртом сразу оба соска и сосет молоко из обоих.
К роду гелад относится один вид Т. gelada с 2 подвидами. Как правило, гелады держатся очень крупными стадами, численность которых может достигать 300—400 особей. Практически гелады — чисто наземные животные и
никогда не карабкаются по деревьям. Питаются они травой, насекомыми, молодыми побегами диких и культурных злаков и их зернами. По жестам в мимике гелады напоминают павианов.
Род мартышек (Cercopithecus) — самый многочисленный по количеству видов род приматов, В настоящее время к нему относят 23 вида с 71 подвидом, многие из которых различными авторами возводились ранее в ранг самостоятельного вида. Мартышек подразделяют на 3 под-рода. ,
В подроде обыкновенных мартышек (Cercopithecus) выделяют 9 групп, включающих 21 вид с 67 подвидами. Они широко расселены в Африке от Сенегала на западе, до Эфиопии и Судана на востоке, а к югу до Анголы.
Обыкновенные мартышки — обезьяны средних раз-меров, массой тела 3—6 кг в с хвостом (55—100 см) более длинным, чем голова и туловище (40—65 см)^ Самки мельче самцов. Шерсть густая, мягкая в короткая; по окраске она сильно варьирует у разных видов, но зеленые, желтые и черные цвета преобладают. Кроме того, для рода характерны яркие локализованные.пятна, полосы, усы и борода. Мордочка умеренной длины. Защечные мешки и седалищные мозоли небольшие. Передние конечности короче задних. Пальцы на кисти длинные с хорошо развитым большим пальцем.
Мартышки встречаются главным образом в дождевых и горных (до 3000 м над уровнем моря) лесах. Очень быстрые в движениях, они хорошо бегают по земле. Возможно, некоторые виды умеют плавать. Снят в густой листве сидя. Питаются листьями, молодыми побегами, плодами, часто совершают набеги на плантации. '
Держатся мартышки стадами в 40—50 особей, но состав и размер стада варьирует в различное время дня. Чаще всего в период передвижения в поисках пищи стадо состоит из 10—15 особей, а спят маленькими группами в 3—5 особей с одним взрослым самцом. Из всех марты-
> 127



 шек наиболее известна земная мартышка (С, sabaeus), которая часто встречается в зоопарках, в различного рода питомниках и часто используется в медицинских целях. Земные мартышки живут в горах Эфиопии {1000—2000 м над уровнем моря). Они крупнее домашней кошки. Плечи, спина и хвост покрыты короткой серой шерстью с желтовато-зеленым оттенком, нижние части светлые, «шапочка» на голове зеленая, бакенбарды и усы белые. Кожа на лице черная, на подошвах и ладонях желтая,
шек наиболее известна земная мартышка (С, sabaeus), которая часто встречается в зоопарках, в различного рода питомниках и часто используется в медицинских целях. Земные мартышки живут в горах Эфиопии {1000—2000 м над уровнем моря). Они крупнее домашней кошки. Плечи, спина и хвост покрыты короткой серой шерстью с желтовато-зеленым оттенком, нижние части светлые, «шапочка» на голове зеленая, бакенбарды и усы белые. Кожа на лице черная, на подошвах и ладонях желтая,
У мартышек стада менее организованы, чем у павианов, менее выражены, звуковая сигнализация более слабая, лицо мартышек менее подвижное и мимика беднее.
К роду красных мартышек гусаров (Erythrocebus) от
носятся наземные обезьяны сухих травянистых и полупу
стынных зон Африки, расположенных к югу от Сахары и
к северу от тропических дождевых лесов. В этот род вклю
чен один вид — обыкновенная мартышка гусар (Е. patas) с
четырьмя локальными подвидами. ,
Будучи наземными животными, гусары хорошо приспособлены к беганию на четырех конечностях. Но они также могут карабкаться по скалам.
Гусары встречаются небольшими группами, состоящими из взрослого самца, нескольких самок и детенышей разных поколений. Самец всегда находится во главе стада и несет сторожевые функции. Пища гусаров разнообразна — трава, плоды деревьев, семена, насекомые, ящерицы, птицы и их яйца, мед.
Подсемейство тонкотелых обезьян (Colobinae) объединяет животных, характеризующихся стройным телом и очень длинным хвостом. Из 6 родов только толстотелы обитают в Африке, остальные — в Юго-Восточной Азии.
Род обыкновеных тпонкотелов, или лангуров (Presbytis), распространен почти по всей Индс-Малайской области. У обыкновенных тонкотелов округлая голова и укороченная морда, длинные стройные конечности. Кисти и особенно стопы очень удлиненные; большой палец
кисти короткий, но хорошо противопоставляется остальным. Хвост (50—110 см) длиннее головы и туловища (41— 78 см), вместе взятых. Это животное средних и крупных размеров, массой тела от 4 до 20 кг. Желудок сложный, состоит из трех отделов. Имеется воздушный горловой мешок. У самцов крупные клыки, у самок они значительно меньше. Шерсть длинная; на голове волосы образуют хохолок или шапочку. Как правило, лицо, ладони и подошвы черные. Цвет шерсти варьирует у разных видов -т-черный, серый, коричневый, золотистый. С возрастом окраска животных сильно меняется.
Обыкновенные тонкотелы, кроме плодов, поедают в большом количестве листья. В отличие от мартышек, это мрачные, тихие животные. В неволе они плохо приспосабливаются и смертность их велика. Но некоторые тонкотелы живут в зоологических садах до 20 лет.
Род гималайских носатых тонкотелов, шшринопи-теков (Rhinopithecus), известен из Северного Вьетнама, Южного и Западного Китая, возможно, встречается в Тибете. Ринопитеков называют также курносыми тонкотелыми обезьянами за короткий вздернутый нос, который у старых особей иногда сильно загибается кверху и почти достигает лба. Тело их плотное с относительно короткими ногами. Длина хвоста равна 61—92 см, головы и туловища — 56—83 см. Шерсть длинная и густая, с возрастом становится длиннее, гуще и ярче окрашенной. Лицо вокруг глаз и носа голое, бледного зелено-голубого цвета.
Род пигатриксов (Pygatnrix) распространен в тропи-;. ческих дождевых лесах Лаоса, Вьетнама и острова Хай-
. Эти обезьяны довольно плотного, коренастого те-гзюсложения, с длиной головы и туловища 55—82 см; хвост почти такой же длины (60—77 см). Самки, как у боль-; шинства обезьян, мельче самцов. Передние конечности
i короче задних. Мех короткий.
В роде носачей обыкновенных (Nasalis) один вид — : носач обыкновенный, или кахау (N. larvatus), с острова
У5. Зах. 466
Носачи — древесные животные. Они живут в лесах и мангровых зарослях вдоль берегов рек и заливов. Обычно держатся на крупных ветвях, прекрасно… Род короткохвостых носатых тонкотелов, или си-миасов (Simias), также включает… Фоц, толстотелое, или гверец (Colobus), — африканские представители тонкотелых обезьян. Они широко расселены в…Семейство гиббоновые (Hylobatidae)
большой палец крупный и способен к схватыванию при лазании. На больших пальцах кистей и стоп ногти плоские и широкие, на остальных — узкие… Шерсть длинная, густая, по цвету варьирует от черной или темно-коричневой до… Белорукий гиббон (Н. 1аг) обитает в Индокитае, Таиланде, на Суматре, на полуострове Малакка. Его белые кисти и стопы…Семейство понгиды (Pongidae)
Род орангутанов (Pongo), как и гиббонов, — азиатская форма антропоидов. К нему относится один вид — обыкновенный орангутан (P. pygmaeus) с двумя… Орангутаны—крупные обезьяны с ярко выраженным половым диморфизмом. При массе… % 135Семейство люди (Hominidae)
v 149 2подвида— неандерталец (H.s. neanderthalensis) и современный человек (H.s.…Глава 6. ИСКОПАЕМЫЕ ПРИМАТЫ
Древнейшие примитивные приматы возникли, очевидно, ещев конце мелового периода. В эту эпоху горообразовательные процессы, сопровождающиеся общим поднятием суши, привели к сокращению площади и обмелению морей в северном полушарии. В южном полушарии, наоборот, сокращалась площадь сущи вследствие разрушения остатков Годваны.
Предками приматов, как и других плацентарных млекопитающих были, по-видимому, древнейшие из плацентарных насекомоядных — эндотерии, остатки одного из представителей которых были найдены в верхнеюрских и нижнемеловых отложениях в горах Лаоса, Северо-Восточном Китае. Эндотерии, как и другие насекомоядные, сформировались, вероятно, в одной из частей Гондваны, скорее всего, в лемурийской ее части, ныне занятой Индийским океаном. Возникшие в конце палеозоя примитивные млекопитающие — предки насекомоядных — могли с успехом выжить в условиях господства пресмыкающихся среди животных и голосеменных среди растений, только используя менее всего употреблявшиеся пресмыкающимися источники питания в виде насекомых. Отсюда и ранняя крайняя специализация некоторых древнейших млекопитающих.





 Так как до распростргийенияпокрьпххеменных растений ландшафт южных материков Гондваны представлял собой чередование открытых пространств и разреженных лесов, большинство древних насекомоядных были наземные животные. ЛшШШШшще мезозоя, когда возникли огромные массивы лесов покрытосемянных, часть насекомоядных, не успевшая специализироваться в поисках и погоне за насекомыми, постепенно стала приспосабливаться к древесному образу жизни. В связи с новым образом жизни на кустарниках и деревьях тело этих насекомоядных должно было претерпеть значительные изменения. В поисках пищи, в погоне за насекомыми им приходилось передвигаться не только по стволам и толстым веткам деревьев, но и по более тонким ветвям, перескакивать с ветки на ветку, с дерева на дерево, лазать по лианам и воздушным растениям (эпифитам). Поэтому прежде всего изменилось строение конечностей этих насекомоядньн.
Так как до распростргийенияпокрьпххеменных растений ландшафт южных материков Гондваны представлял собой чередование открытых пространств и разреженных лесов, большинство древних насекомоядных были наземные животные. ЛшШШШшще мезозоя, когда возникли огромные массивы лесов покрытосемянных, часть насекомоядных, не успевшая специализироваться в поисках и погоне за насекомыми, постепенно стала приспосабливаться к древесному образу жизни. В связи с новым образом жизни на кустарниках и деревьях тело этих насекомоядных должно было претерпеть значительные изменения. В поисках пищи, в погоне за насекомыми им приходилось передвигаться не только по стволам и толстым веткам деревьев, но и по более тонким ветвям, перескакивать с ветки на ветку, с дерева на дерево, лазать по лианам и воздушным растениям (эпифитам). Поэтому прежде всего изменилось строение конечностей этих насекомоядньн.
Некоторые насекомоядные приспособились к передви
жению прыжками; Это повлекло за собой развитие зад
них конечностей, которые постепенно удлинились и ста
ли более мощными, чем передние, и у них должны были
развиваться цепляющие функции. Когти на пальцах зад
них ног, поскольку они не были втягивающимися, как у
кошачьих, становились менее полезными: постепенно уко
рачиваясь и уплощаясь, они превратились в ногти. На паль
цах передних конечностей у таких насекомоядных сохра
нились когти, способствуя фиксации за ту ветку, на кото
рую совершается прыжок. Подобного рода специализация
конечностей отмечается, например, у примитивного при
мата анагале гобийского (Anagale gobiensis) из олигоцевд-
вых отложений Монголии: его считают родственников
тупайи. .
У других древних насекомоядных специализация в прыжках помимо соответствующего изменения задних конечностей в целом вызвала также и удлинение пяточного
отдела стопы, так что их задние конечности стали представлять собой двойной рычаг, позволявший им, несмотря на небольшие размеры тела, совершать огромные прыжки. В этом случае для фиксации за ту ветвь, на которую зверек прыгал, одних когтей оказалось недостаточно, и передние конечности должны были развить способность к захвату ветвей. Это привело к удлинению пальцев передней конечности и усилению возможности раздельного манипулирования ими. Необходимость лазания по деревьям привела и к возникновению способности большого пальца кисти противопоставляться остальным, что позволило охватить тонкие ветки. Так сформировались представители подотряда долгопятовых, или терзиоидов.
Способность держать тело в вертикальном положении развивалась у предков приматов также под воздействием их образа жизни: чем большее значение приобретало лазанье по стволам и лианам, тем чаще тело принимало вертикальное положение, а это вело к укреплению позвоночного столба, перестройке мышц спины, изменению функций передних и задних конечностей. Очевидно, предки долгопятовых чаще, чем предки тупаий, пользовались таким способом лазанья, поэтому у них развивались вертикальная постановка тела и хватательная способность передних конечностей. Отчасти следствием этого было и то, что длина морды укорачивалась, а череп становился более округлым, чем у типичных четвероногих животных.
Перескакивание с ветки на ветку и с дерева на дерево вело также к изменению органов зрения. Необходимость • точной ориентировки и оценки расстояния при передвижении привела к возникновению стереоскопичности: зрительные оси обоих глаз должны были получить большее хождение, что могло быть достигнуто перемещением глаз 5 вперед, на лицевую сторону черепа. Такого изменения не Й>ыло у предковых форм приматов, нет и у современных
| V |






 тупаий, на осуществилось в ходе эволюции у долгопятовых. Правда, долгопятовые отличаются огромными размерами глаз, но эта их особенность возникла вследствие ночного образа жизнн. В соответствии с прогрессом органов зрения, к которым перешла особо важная роль в передвижении и ориентировке в обстановке, обоняние у приматов по сравнению с наземными животными несколько ослабилось.
тупаий, на осуществилось в ходе эволюции у долгопятовых. Правда, долгопятовые отличаются огромными размерами глаз, но эта их особенность возникла вследствие ночного образа жизнн. В соответствии с прогрессом органов зрения, к которым перешла особо важная роль в передвижении и ориентировке в обстановке, обоняние у приматов по сравнению с наземными животными несколько ослабилось.
Третья группа древнейших приматов, выделившаяся, по-видимому, одновременно с долгопятовыми, приспособилась преимущественно к лазанью. Поэтому передние и задние конечности у них специализировались как хватательные. Кисти и стопы их должны были действовать подобно клещам, зажимая ветви или лианы между большим и остальными пальцами. В этом случае не исключалось также передвижение прыжками, хотя такой способ передвижения играл второстепенную роль. При лазании вертикальное положение телабылоне менее обычным, чем горизонтальное или наклонное, и у потомков третьей группы примитивных приматов с течением времени начали накапливаться черты, все более и более приближавшиеся к обезьяньим. Кроме того, хватательные способности конечностей привели к образованию специальных приемников осязательных раздражений в виде выпуклых осязательных подушечек, покрытых капиллярными линиями и узорами. У обезьян и человека подобными узорами сплошь покрыта кожа ладоней и подошва ступней. Особая сложность и разнообразие движений в сочетании с сильной подвижностью свойственны большинству древесных млекопитающих, привели к значительному развитию моторных участков мозга. Переход лазающих древнейших приматов, питавшихся насекомыми, к употреблению растительной пищи обогащал их организм более разнообразными веществами. Это тоже стимулировало усложнение и рост объема мозга, который и у низших приматов развит довольно сильно.
Эта третья группа животных, переходных от насекомоядных к приматам, и дала в начале палеогена ветвь полуобезьян, или лемуров. Именно эти низшие приматы стали употреблять в пищу помимо насекомых ягоды, плоды и другие съедобные части растений. Изменение пищи и увеличение ее разнообразия вызвали прежде всего преобразование челюстей и зубов. Часть зубов оказалась лишней и исчезла, другие претерпели изменение формы и структуры, становились способными перетирать любую пищу. Так, если у древних насекомоядных было 44 зуба — столько же, сколько установлено у анагале гобийского, то у ископаемых долгопятовых их количество сокращается до 40—28 (у современных долгопятов — 34), а у лемуров до40—32.
Низшие приматы
По этому представлению предки тупайи вели полудревесный образ жизни. Хвост бьш длинный, конечности сравнительно короткие, пятипалые с когтями; в… 155 матичность), но уже с признаками начинающегося развития зрительной области. Мордочка длинная, ноздри на конце…Ископаемые тупайи
Ископаемые лемуры
Семейство адапид (Adapidae): зубная формула 214 3/ 2 14 3 (молочные 2 1 3/2 1 3), резцы стоят более или менее вертикально, невелики, клыки умеренных… Череп адапид низкий и широкий с сагиттальным и затылочным гребнями, суживается… '* 157 .Ископаемые долгопяты
паемые долгопяты объединяются в семейство анаптомор-фид (Anaptomorphidae Соре). Их общая характеристика следующая: зубная формула 2 1 3 3/2—0 1 3—2… Известно 13 родов североамериканских ископаемых форм долгопятовых, из которых… Анаптоморф эмулюс (Anaptomophus aemulus), известный по нижней челюсти из эоцена Бриджера, Уайоминг, отличается от…Зак. 486
Основная форма европейских долгопятов — некроле- мур (Necrolemur Filhal), от которого известны 6 черепов, бедро и большеберцовая кость, на… Микрохерус (Microchoerus Wood) — вид, родственный некролемуру, из эоцена… Периконодон (Periconodon Stehtin) — вид европейских ископаемых долгопят из эоцена Швейцарии. Характерной особенностью…Высшие приматы
История эволюции отряда приматов падает в основном на третичный период, имеющий следующие подразделения: палеоцен, эоцен, олигоцен, миоцен,…Ископаемые широконосые обезьяны
(Aotinae)» Для американских обезьян исходными формами развития служили… торых одной из самых примитивных форм является омо-мис. Проникновение предков этих обезьян в Южную Америку должно…Низшие узконосые обезьяны
Ореопитек (Oreopithecus bambolii) известен по фрагментам нижней и верхней челюстей из нижнего плиоцена Ь' Тосканы (Италия) и Бессарабии. Зубная формула 2 12 3/ 212 3, как и у прочих… Прочие ископаемые мартышковые обезьяны относятся к макакам и павианам, некогда более широко распространенным по…Ископаемые человекообразные обезьяны
Древнейшим и самым примитивным их представителем является парапитек (Parapithecus ftaasi) из нижнего олигоцена Файюма (Египет). От него известна… В нижнем олигоцене Египта была найдена нижняя челюсть и другой обезьяны. В… Примерно с 1962 г. Элвин Саймоне производил раскопки в Файюме (Египет), где нашел новые остатки проплиопитека, а…Рис. 38. Орудия олдувайской эпохи (по Campbell, 198S)
'*■ 181

| 3,5 |
 Таким образом, на протяжении примерно 2800 км с севера на юг на Африканском материке в период от 1 до 4 миллионов лет обитали существа, которые по способу своего передвижения (двуногости) и по строению зубной системы были гораздо ближе к людям, чем к антропоморфным обезьянам. Однако по форме эндокрана они были более сходны с шимпанзе, чем с человеком. По абсолютному объему головного мозга они также значительно уступали людям и в целом лишь' немного превосходили шимпанзе и гориллу. Но следует учитывать, что по массе тела они были в общем гораздо легче гориллы и даже шимпанзе. Их стопа была весьма сходной с человеческой, а кисть еще обладала довольно многими архаическими чертами. Значение этих находок в том, что они заполняют пробел в цепи существ, ведущей от животных к людям, и увеличивают аргументы в пользу признания Африки прародиной человечества.
Таким образом, на протяжении примерно 2800 км с севера на юг на Африканском материке в период от 1 до 4 миллионов лет обитали существа, которые по способу своего передвижения (двуногости) и по строению зубной системы были гораздо ближе к людям, чем к антропоморфным обезьянам. Однако по форме эндокрана они были более сходны с шимпанзе, чем с человеком. По абсолютному объему головного мозга они также значительно уступали людям и в целом лишь' немного превосходили шимпанзе и гориллу. Но следует учитывать, что по массе тела они были в общем гораздо легче гориллы и даже шимпанзе. Их стопа была весьма сходной с человеческой, а кисть еще обладала довольно многими архаическими чертами. Значение этих находок в том, что они заполняют пробел в цепи существ, ведущей от животных к людям, и увеличивают аргументы в пользу признания Африки прародиной человечества.
На основании данных палеогеографии (саванны, полупустыни) и морфологии австралолитековых, а также того обстоятельства, что некоторые из них умели изготавливать простейшие орудия, большинство антропологов мира включают австралопитеков в семейство гоминид в качестве подсемейства австралопитековых (рис. 39).
Ископаемые гоминиды
4.0
Рис.39. Родословное дерево гоминид <по Джохансон, Иди, 1984)
Кроме найденных остатков Дюбуа еще в ноябре 1890 г. обнаружил у р. Брубус, в 40 км к востоку от Три-ниля, фрагмент нижней челюсти человеческого типа с сохранившейся ячейкой клыка и двумя предкоренными
'v 183

 зубами; подбородочный отдел не сохранился. Челюсть была найдена в том же геологическом слое и с той же ископаемой фауной (стегодон и другие исчезнувшие млекопитающие), что и кости питекантропа; степень минерализации также была сходной. Эта челюсть, низкая и относительно толстая, отличалась замечательной особенностью — огромной Протяженностью в ширину ямки для двубрюшной мышцы (16 мм). Такие большие ямки не свойственны ни обезьянам, ни людям, у которых этот размер варьирует от 5,0 до 7,5 мм.
зубами; подбородочный отдел не сохранился. Челюсть была найдена в том же геологическом слое и с той же ископаемой фауной (стегодон и другие исчезнувшие млекопитающие), что и кости питекантропа; степень минерализации также была сходной. Эта челюсть, низкая и относительно толстая, отличалась замечательной особенностью — огромной Протяженностью в ширину ямки для двубрюшной мышцы (16 мм). Такие большие ямки не свойственны ни обезьянам, ни людям, у которых этот размер варьирует от 5,0 до 7,5 мм.
Найденные вместе с черепной крышкой зубы питекантропа различны по своему типу: коренные, по-видимому, имеют черты сходства с орангутаном, предкорен-ной очень похож на современный человеческий. Наиболее важная особенность черепной крышки—ее промежуточное по форме и по величине положение между человеком и антропоморфной обезьяной. Так, при реконструкции недостающих частей емкость полости мозгового отдела черепа оказалась равной приблиз^ельно 900 см3, т.е. величине, лежащей примерно посередине между средней емкостью черепа гориллы (500 см3) и современного человека (1400 см3). Лоб у питекантропа покатый, весьма приближающийся в этом отношении к шимпанзе. Свод черепа очень низкий; его затылочный отдел уплощен сверху. Наибольшая ширина черепа расположена низко, ближе к основанию. При рассмотрении черепа сверху можно видеть, что его наибольшая ширина сдвинута к заднему отделу, как у шимпанзе, в отличие от Человека, у которого наибольшая ширина расположена примерно посредине. На любой кости выделяется надглазничный валик, имеющий форму навеса. По средней линии лобной кости идет вали-кообразное возвышение, заканчивающееся пологим бугром в области венечного шва.
Слепок черепной полости питекантропа позволяет сделать выводы о некоторых особенностях формы его головного мозга. По своей структуре он значительно ближе к
человеческому, чем к мозгу любой антропоморфной обезьяны, но обладает, однако, весьма примитивными чертами. Так, нижняя лобная извилина у него менее развита, чем у современного человека. «Лобная крышка» расположена выше, чем у современных людей, вследствие чего глазничная поверхность лобной доли более открыта—чер-(та, которая еще резче выражена у высших обезьян. Теменная доля мозга также менее развита, чем у человека.
Бедренная кость по размерам и по строению очень напоминает человеческую, ее длина 455 мм (у современного человека такая длина бедра соответствует длине тела, приблизительно в 165—170 см).
Находка питекантропа сыграла исключительную роль в антропологии. Хотя ранее были известны ископаемые черепа и фрагменты скелета неандертальского человека, однако они были менее примитивны по своей организации, чем питекантроп, а главное — не отличались от современного человека по объему мозга. Вследствие этого именно яванская находка была оценена как важнейшее доказательство правильности теории Дарвина о происхождении человека от высшей обезьяны. .
Кроме этих находок, в общем близких по своему типу к тому представлению о питекантропе, которое сложилось у наиболее компетентных исследователей, были сделаны еще две находки, заслуживающие особого рассмотрения, так как принадлежность их к группе питекантропов сомнительна (фрагменты нижней челюсти), получившей, благодаря своим размерам, название мегантропа.
При изучении известняковых пещер вблизи Пекина в начале 20-х гг. XX в. было обнаружено много ископаемых остатков животных и среди них два зуба, принадлежащих примитивному обезьяночеловеку среднеплейсто-ценового периода, давностью примерно 500 тыс. лет назад. Открывший их Д. Блэк дал им название Sinanthropus pekmensis. При дальнейших раскопках в этих пещерах были
ч 185

 обнаружены части скелетов более чем 40 особей синантропов — мужчин и женщин, старых и молодых.
обнаружены части скелетов более чем 40 особей синантропов — мужчин и женщин, старых и молодых.
Череп синантропа с массивными надглазничными валиками и низким, покатым лбом очень похож на череп питекантропа. Челюсти у синантропа массивные, лишенные подбородка, весьма сходные с челюстями обезьян, нос широкий и плоский (рис. 40).
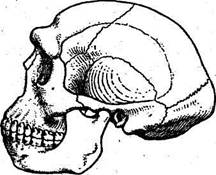
Рис. 40. Череп синантропа
Полость мозгового черепа была в среднем объемистее, чем у питекантропа: от 850 см3 у одного из женских черепов до 1220 у одного из мужских. По возвышенности теменной области мозг синантропа обнаруживал переход к неандертальцам, но заостренность и обращенность вниз лобных долей делала его сходным с мозгом африканских антропоидов.
К примитивным особенностям лицевого скелета синантропа нужно отнести сильное выступание верхних челюстей (прогнатизм). В связи с тем, что мозговой отдел черепа по сравнению с лицевым очень мал, для поддержания головы в равновесии необходимо было сильное развитие мышц затылка, а следовательно, и увеличение площади выйной области затылочной кости. Если принять за
100 расстояние от точки ламбда до точки инион (т.е. длину верхней половины чешуи затылочной кости), то размер выйной области будет равен у синантропа 123, а у современного человека в среднем только 62. С укреплением шейной мускулатуры у синантропа связано также очень сильное развитие затылочного валика.
Вместе с тем сам факт изготовления синантропом орудий доказывает, что руки его были свободны, т.е., что он передвигался на двух ногах. Один из фрагментов бедренной кости позволяет охарактеризовать ее как почти пря-
I мую, массивную, с хорошо выраженной шероховатой линией. Восстанавливая приближенно полную длину бедренных костей, можцо предположить, что рост мужской особи синантропа равнялся 162 см, а женской — 152 см.
К числу наиболее важных находок ископаемого человека принадлежит нижняя челюсть с зубами (рис. 41), обнаруженная в 1907 г. близ города Гейдельберга немецким антропологом О. Шетензаком, который дал наименование находке — гейдельбергский человек. Своеобразные
|
| Рис. 41. Нижняя челюсть гейдельбергского человека |
| черты челюсти гейдельбергского человека заключаются в следующем: вырезка восходящей ветви очень мала, венечный отросток широк и закруглен; сочленовная головка суставного отростка весьма велика и отличается по форме: у современного человека сочленовные головки имеют


 вид узких, поперечно направленных валиков, приспособленных главным образом к шарнирным движениям в сагиттальной плоскости. У гейдельбергского человека головки очень широки в мезодистальном направлении, по форме далеки от цилиндра, что, по-видимому, указывает на иной характер жевательных движений. Подбородочные отверстия велики, имеются маленькие дополнительные отверстия. Нижний контур симфиза вогнут, так что при рассмотрении челюсти спереди передний край ее заметно приподнят над горизонтальной плоскостью. Однако весь зубной ряд и каждый зуб в отдельности почти не отличаются от зубов современных людей: клыки не имеют конической формы, не выдаются над общим рядом зубов, диа-стемы отсутствуют.
вид узких, поперечно направленных валиков, приспособленных главным образом к шарнирным движениям в сагиттальной плоскости. У гейдельбергского человека головки очень широки в мезодистальном направлении, по форме далеки от цилиндра, что, по-видимому, указывает на иной характер жевательных движений. Подбородочные отверстия велики, имеются маленькие дополнительные отверстия. Нижний контур симфиза вогнут, так что при рассмотрении челюсти спереди передний край ее заметно приподнят над горизонтальной плоскостью. Однако весь зубной ряд и каждый зуб в отдельности почти не отличаются от зубов современных людей: клыки не имеют конической формы, не выдаются над общим рядом зубов, диа-стемы отсутствуют.
Питекантропы, синантропы, гейдельбергский человек
и другие объединяются под названием «архантропы»
(archaios —старый -t anthropos — человек). Костные ос
татки архантропов известны из раннего и среднего плей
стоцена Азии, Африки и Европы (абс. возраст—от 1,9 млн
лет до 360 тыс. лет). Эти представители ископаемых го-
минид относятся к виду Homo erectus - человек выпрям
ленный. Архантропы характеризовались высоким надбро
вьем и мощным надглазничным валиком, покатым лбом,
низким сводом черепа с уплощением его затылочного от
дела. По средней линии лобной кости имеется валикооб-
разное возвышение, отмечены также сильный прогнатизм,
отсутствие подбородочного выступа нижней челюсти. Эн-
докраны свидетельствуют о разрастании корковых полей,
регулирующих направленные движения рук, развитие зон,
которые обеспечивают анализ сигналов, поступающих от
зрительных, слуховых и тактильных центров. Эти особен
ности мозга позволяли совершенствовать и звуковую сиг
нализацию. -
Способность к членораздельной речи зависит от таких структурных особенностей, как загиб корня языка в гортанную полость, усиление голосовых связок, разрастание внутренних краев черпаловидных хрящей гертани,
появление которых привело к более четкой дифференциации издаваемых звуков благодаря разделению выдыхаемого воздуха на верхнюю струю, или носовую, и нижнюю, или ротовую. Увеличение подвижности языка способствовало четкости произнесения качественно разнообразных звуковХгУбные' нёбные, зубные). Но массивные нижние челюсти архантропов препятствовали быстрой смене артикуляции и плавности речи, которая совершенствовалась не только по мере развития абстрактного мышления, но и постепенного морфологического преобразования лицевого скелета.
Этот этап связан с зарождением абстрактного мышления и развитием общественных отношений.-У архантропов фиксируются не только устойчивые типы орудий, но и локальные особенности каменной индустрии: преимущественное использование чопперов (галечных орудий со стесанными краями или верхушками), отщепов с зубчатыми краями на одних стоянках и рубил, колунов — на других, что связывают с природными условиями и характером сырья, используемого для изготовления орудий. Изучение экологического окружения показало, что расселение архантропов происходило на фоне резких изменений климата — из-за смены ледниковых периодов. В трудных климатических условиях применение разнообразных орудий труда, использование огня и усовершенствование жилища позволяли долгое время жить на одном и том же месте или возвращаться к месту обитания после длительных переходов. Эти первобытные коллективы были невелики. По данным палеодемографических исследований, у синантропов они состояли из 3—6 особей мужского пола и 6—10 женских особей и 15—20 детей. В борьбе за выживание самым главным было тесное сплочение первобытной общины, в чем немаловажную роль играло наличие у архантропов зачатков членораздельной речи.
Следующий этап эволюции связан с древними людьми — палеоантропами (palaios — древний + anthropos — человек). Палеоантропы — это обобщенное название ис-
*■ 189
копаемых людей, которых рассматривают как вторую ста
дию эволюции человека, следующую за архантропами и
предшествующую неантропам. Костные остатки палеоан
тропов известны из среднего и позднего плейстоцена Ев
ропы, Азии и Африки. Абсолютный возраст от.-250 до
40 тыс. лет. Первая находка черепа палеоантропа была сде
лана в 1848 г. в Гибралтарской крепости. И хотя она по
ступила в распоряжение ученых, о ней вспомнили лишь
после открытия в 1856 г. черепа и костных остатков в до
лине реки Неандерталь близ Дюссельдорфа, в связи с чем
этот вид палеоантропов был назван неандертальским че
ловеком. '
По времени существования и морфологическим особенностям выделяют 2 группы неандертальцев. Ранние неандертальцы жили примерно 150 тыс. лет назад. Для них характерны: вертикальный профиль лица, несколько редуцированный надбровный рельеф, более прогрессивное строение зубов (рис. 42). Объем мозга варьировал от 1200

Рис. 42. Черепа неандертальцев (по Вгокеп-НШ, 1921): а — из Chapelle-aux-Samts; б— из Rhodesia
11400 см3. Древность более поздних неандертальцев достигает 80—35 тыс. лет. Характерными особенностями этих гоминид является сильное развитие надбровья, сжатый сверху вниз затылочный отдел, широкое носовое отвер-стие, наличие затылочного валика и другие черты специализации. Особенно сильно специализация затронула верхнюю конечность, на что указывает отсутствие седловид-вости первого запястно-пястного сустава, свидетельству- Ющее об ограничении подвижности и манипуляционной ^пос°бности и усилении силового захвата. У некоторых поздних «классических» неандертальцев — палестинские ^Ваходки и гоминиды Староселья (Крым), Тешик-Таш (Узбекистан) — много признаков близости к современным
ЛЮДЯМ.
В культуре неандертальцев появляется большое количество новых элементов по сравнению с предыдущими стадиями. С неандертальским типом синхронен мустьер-ский этап в развитии палеолитической культуры (назван по пещере Ле Мустье в Юго-Западной Франции), датируемый 100—40 тысячелетиями. В инвентаре многих мусть-ерских стоянок сохранилась традиция двусторонней обработки камня. Так, например, во многих стоянках Крыма найдено большое количество двусторонне обработанных рубилец, по форме напоминающих шелльские и ашельс-кие, но значительно более миниатюрных. Господствующее положение занимают скребло и остроконечник. Кремниевые отщепы откалывались от дисковидного нуклеуса (ядршца).
Способ ударной ретуши ашельской эпохи в мустьер-скую эпоху получил дальнейшее развитие — была изобретена контрударная ретушь. Новый способ заключался в том, что обрабатываемое орудие помещали на каменную или костяную подставкух(наковальню) и ударяли по нему деревянной колотушкой. Скребло обрабатывалось лишь по одному краю, который называется режущим, и предназначалось, по-видимому, для выскабливания шкур. Ос-

 троконечник обрабатывался с двух сторонщ использовался в качестве ударного и режущего инструмента. Весьма вероятно, что при этом он прикреплялся* палке, тогда получалось какое-то орудие вроде копья, либо привязывался к короткой рукоятке и употреблялся наподобие ножа. Если это предположение верно, то, значит, уже тогда появились составные орудия. В поздних мустьерских стоянках эти типы орудий дополняются по-разному обработанными удлиненными пластинками. Здесь уже видно зарождение верхнепалеолитической техники.
троконечник обрабатывался с двух сторонщ использовался в качестве ударного и режущего инструмента. Весьма вероятно, что при этом он прикреплялся* палке, тогда получалось какое-то орудие вроде копья, либо привязывался к короткой рукоятке и употреблялся наподобие ножа. Если это предположение верно, то, значит, уже тогда появились составные орудия. В поздних мустьерских стоянках эти типы орудий дополняются по-разному обработанными удлиненными пластинками. Здесь уже видно зарождение верхнепалеолитической техники.
Охота давала неандертальцам обильный материал для костяных поделок. Началось использование кости для производственных целей (наковаленки, ретушеры, острия), для изготовления мелких заостренных орудий. Обнаруженные на многих стоянках архантропов и на всех у неандертальцев следы кострищ, а в ряде случаев и преемственность в использовании огня архантропами и неандертальцами позволяют предположить, что отдельные группы древнейших людей научились добывать огонь. В последующем добывание и использование огня в быту явилось одним из элементов разнообразной по содержанию культуры неандертальца. Присущие ему усовершенствованные орудия охоты и обработки кожи, способность сооружать сложные жилища имеют давние традиции.
У палеоантропов формируется абстрактное мышление, зарождается искусство и общественное сознание. Это подтверждают захоронения умерших, выполненные по определенному обряду: Об уровне развития мышления, альтруизме и зарождении идеологических воззрений неандертальцев ярко свидетельствуют находки. На территории Ирака в пещере Шанидар обнаружены следы кострищ, кремневые мустьерские орудия и скелеты неандертальцев, древность которых составляла 50—60 тыс. лет. Один из мужских скелетов носил на себе следы тяжелых прижизненных повреждений — травмы черепа и ампутации нижней части правой руки. Необычайная стертость зубов ука-
?1^ивает на то, что этот неандерталец пользовался ими при {работе, компенсируя утраченную правую руку. С такими повреждениями он мог оставаться в живых лишь при оказании ему помощи другими членами сообщества. Палео-,,. -ботанические исследования почвы, взятой из этой пещеры, позволили утверждать, что при захоронении неандер- ..телец был уложен на подстилку из веток и покрыт цвета-•-■ ■ Г- ми. О зарождении искусства свидетельствует открытие и ?£| использование естественных красителей. Не совсем чет-|у кие орнаментальные композиции наносились в виде пя-£ тен красок на стены пещеры.
§■ Следующий этап эволюции человека начался около !Г 200тыс. лет назад с возникновения неоантропов, прямых ' i предшественников современного человека. Первая наход-'(, ка неоантропа была сделана в гроте Кро-Маньон во Фран-й' ции, где была открыта многослойная стоянка возрастом около 40 тыс. лет. По имени грота, в котором были найдены ископаемые остатки неоантропов называют кромань-
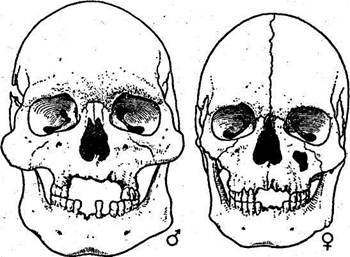
|
| Рве. 43. Черепа кроманьонцев % 193 |
| 7. Зак. 486 |
t
сипами. Стоянки неоантропов, живших около Звтыс. лег назад, были обнаружены в Белоруссии, на Украине, в России, на Кавказе, Памире, в других районах Средней Азии, в бассейнах рек Камы, Индигирки, Печоры, Десны, на Камчатке. В этот период человек расселялся по всей суше, осваивая различные природные области. По сравнению с неандертальцами человек современного вида выделяется прогрессивными особенностями: более сильным развитием височной и лобной долей мозга, что свидетельствует о совершенстве мозговой деятельности. Произошли и структурные изменения черепа: отсутствует агаесивный надглазничный валик, выступает подбородок, об увеличении лоб-ных долей мозга, связанных с умственной деятельностью, свидетельствуетвысокий лоб (рис 43). Отныне живущих людей неоантропов отличает общая массивность скелета. Отдельные находки имеют особенности, которые показывают, что все формы неоантропов с самого начала своего существования по морфологическому облику распадались на ряд расовых вариантов. Это важно для понимания процессов расообразования. Так, черепа немолодой женщины и юноши, обнаруженные в гроте Гримальди близ Ментоны (Италия), резко отличались от типичных кроманьонцев, найденных в том же гроте, мозаичным сочетанием расовых черт, совмещая в себе европеоидные и негроидные черты (выступающие челюсти — прогнатизм и широкий, слабовыступающий нос). Такие же черты прослежены и у отдельных кроманьонцев на территории России. На стоянке близ д. Костенки Воронежской области открыто несколько погребений, в которых .обнаружены скелеты детей и взрослых. У самой древней находки (около 30 тыс. лет назад) — скелет мужчины 20—25 лет из захоронения Маркина Горка — при таких европеоидных чертах, как сильное выступание носа, отмечены характерные для негроидов черты: прогнатизм и широкий нос. Значительный интерес с точки зрения локальных особенностей типа и культуры представляет стоянка Сунгирь
адимирской области, открытая в 1956 г. Древность сто-
f явки около 25 тыс. лет. Люди из Сунгиря жили в суровых климатических условиях приледниковья. Судя по инвентарю, их основным занятием была охота. В захоронениях сохранились скелеты мужчины 60 лет и двух детей — мальчика 12-^-14 лет и девочки 9—-11 лет. По морфологи-ческим особенностям это люди современного вида, сходные с кроманьонцами Западной Европы. По несколько уплощенному лицевому скелету и носовым костям можно говорить о некотором налете монголоидности или зарождении этих черт. Рост сунгирца был 180 см, при выраженной массивности скелета он обладал значительной шириной плеч. Прослеживаются различия и в культуре: отдельные локальные варианты культуры можно выделить в каждом районе. Они связаны с типом обработки орудия — камень расщепляется ударом или отжимом — и преимущественным распространением тех или иных орудий, что в свою очередь связано с типом хозяйства. Так, у племен охотников распространены остроконечники, сверла, резцы, наконечники копий и другие орудия из камня и кости. Племена, занятием которых была рыбная ловля, пользовались преимущественно гарпунами, другие виды орудий носили вспомогательный характер. Уже в период палеолита неоантроп из камня и кости изготовлял весьма совершенные орудия труда„ умел рисовать.
Сравнивая морфологические особенности предшественников человека и представителей семейства гоминид, можно выделить основную особенность эволюции — усиление характерного для человека комплекса признаков, так называемой гоминидной триады: прямохождения, манипулятор ных особенностей строения руки, нарастания , массы мозга и усложнения его структуры и функций. Приспособление к прямохождению привело к структурным преобразованиям обеих конечностей, позвоночного столба, тазового пояса, грудной клетки. Изменились пропорции тела. Освобождение верхних конечностей расширило
7* г, 195

 возможности манипуляторной деятельности. Зарождение трудовых операций сопровождалось постепенной анато-f мической перестройкой верхней конечности, особенно кисти, в Направлении обеспечения большей подвижности и прочности суставного аппарата — способности к захвату. Сильное развитие головного мозга, особенно его относительной массы— специфически человеческая черта. По сравнению с обезьянами у человека наблюдается усложнение и дифференциация цитоархитектонических полей, главным образом йередней части лобной и теменной долей. Большая часть коры головного мозга связана с речью. Морфологическая эволюция человека подтверждается косвенными р прямыми доказательствами.
возможности манипуляторной деятельности. Зарождение трудовых операций сопровождалось постепенной анато-f мической перестройкой верхней конечности, особенно кисти, в Направлении обеспечения большей подвижности и прочности суставного аппарата — способности к захвату. Сильное развитие головного мозга, особенно его относительной массы— специфически человеческая черта. По сравнению с обезьянами у человека наблюдается усложнение и дифференциация цитоархитектонических полей, главным образом йередней части лобной и теменной долей. Большая часть коры головного мозга связана с речью. Морфологическая эволюция человека подтверждается косвенными р прямыми доказательствами.
Лава 7. МЕСТО ЧЕЛОВЕКА В ПРИРОДЕ
К середине XIX в. успехи анатомии и других биологических наук подготовили почву для обобщений, которые сделал Дарвин своей теорией эволюции.… * 197 областей естествознания, а также из смежных дисциплин (психологии, истории культуры и др.), на основе которого Дарвин…Черты строения человека, общие с позвоночными
1) у человека есть внутренний скелет, растущий вместе с ростом организма. Он возникает в виде спинной струны, а позднее позвоночного столба,… 2) человек, как и все позвоночные, имеет трубчатое стро ение центральной… 19ЯЧерты строения человека, общие с млекопитающими
Млекопитающие возникли из рептилий в мезозойскую эру, а в настоящее время заняли господствующее положение. Первые млекопитающие были мелкими, мало… Развитие мозга было другим, не менее важным преимуществом млекопитающих. Вся… *■ 201Черты строения человека, общие с приматами
* 203 G большой подвижностью передней конечности свя- F зано строение плечевого… Задняя конечность также приобрела подвижность, соединенную с хватательной способностью, и с внешней стороны стопа…Опорно-двигательный аппарат
1) значительно более короткие фаланги и плюсневые ко сти по отношению к длине предплюсны, чем у выс ших обезьян; в процентах длины стопы длина… 2) очень длинный и очень толстый первый палец по сравнению с другими… 3) значительно меньшая подвижность первого пальца стопы, анатомически выражающаяся в различных осо-Абсолютная и относительная масса мозга у человека и антропоидных обезьян (Рогинский, 1978)
абсолютная относительная Человек 1:45 Шимпанзе 1:61 Горилла … 3) вполне развитая сильвиева щель и полная погружен ность островка Рейлия в ее глубине;Гортань
Для системы мышц гортани человека характерны чет-[ взаимная обособленность отдельных мышц, напри-. мер, боковой перстевидно-черпаловидной от… Для связочного аппарата гортани человека типичны более плотные и более мощные,… Для человека характерно низкое положение гортани: верхний край надгортанного хряща при спокойном положении гортани…Череп и зубы
В соответствии с большой длиной нижней челюсти и большой длиной лицевого отдела в целом, челюсти антропоидов сильно выступают вперед — их черепа… Малые размеры человеческого лица объяснялись разными причинами: общее… f$o стоящего существа, так как чрезмерно большой объем эловы повьппал бы и без того значительную неустойчи-Рвость…Глава 8. ПРОИСХОЖДЕНИЕ ЧЕЛОВЕЧЕСКОГО ОБЩЕСТВА
 | |||
 | |||
 | |||
|

|

|

|
Реконструкция изготовления каменных орудий труда кроманьонцев
При обсуждений проблемы происхождения человеческого общества ученые вынуждены в значительной степени опираться на умозрительные гипотезы и предположения. Эти гипотезы, однако, не всегда беспочвенны, порой они основаны на вполне реальных данных. Наибольшую ценность представляют фактические данные, касающиеся поведения приматов в сообществе, а также австралопитеков и древнейших людей, полученные главным образом при изучении ископаемых остатков. Первая группа данных позволяет более четко представить характер организации сообщества у приматов; вторая дает возможность оценить экологические и физические условия, в которых существовали наши гоминидные предки. Изучая поведение различных видов приматов, можно получить представление о различных формах организации сообщества и можно сопоставить их с учетом особенностей самих приматов и экологических условий, в которых они существуют (см. главу 6), с тем, что известно об условиях жизни наших далеких предков. В этом состоит преиму-щество изучения приматов по сравнению с изучением организации современного человеческого общества. Анализ структуры сообществ приматов имеет большую ценность
Г "9;*ж 486

 для создания моделей организации сообществ догоминид-ных форм приматов, чем изучение ныне живущих примитивных сообществ охотников или собирателей. Действительно, организация сообществ претерпела коренные изменения в ходе эволюции, в основном в результате усложнения структуры мозга и совершенствования речевых навыков. В то же время на примере приматов можно видеть внутригрушювые отношения, основанные на системе сигналов и процессах обучения, не связанных с использованием речи.
для создания моделей организации сообществ догоминид-ных форм приматов, чем изучение ныне живущих примитивных сообществ охотников или собирателей. Действительно, организация сообществ претерпела коренные изменения в ходе эволюции, в основном в результате усложнения структуры мозга и совершенствования речевых навыков. В то же время на примере приматов можно видеть внутригрушювые отношения, основанные на системе сигналов и процессах обучения, не связанных с использованием речи.
Первобытное человеческое стадо
Начальную форму организации общества называют первобытным человеческим стадом. Правда, ряд ученых считает, что употребление этого термина… »развития. Но это возражение вряд ли основательно. Гермин «первобытное… f'■.«.., Эпоха первобытного человеческого стада, очевидно, совпадает с выделением человека из животного мира и…Роль охоты в первобытном стаде
Объектами охоты в зависимости от фауны той или иной области были различные животные. Б тропической ie это были гиппопотамы, тапиры, антилопы, дикие ! и т. д. Иногда среди костей… *■ 229Возникновение общинно-родового
Строя
Крупные сдвиги в развитии производительных сил
У повлекли за собой не менее крупные изменения в организации общества. Возросшая техническая вооруженность человека в его борьбе с природой сделала возможным существование относительно постоянных хозяйственных коллективов. Но в то же время она требовала эффективного использования преемственности и дальнейшего совершенствования усложнившихся орудий и навыков тру-
| да. Первобытному стаду с его относительно аморфной неустойчивой структурой эта задача была не под силу, поэтому стадо неизбежно должно было уступить место более прочной форме общественной организации.
Рядюбстоятельств определял характер этой организации. Во-первых, при крайне низком уровне развития ран-
| непалеолитического общества, в условиях которого начала складываться новая организация, едва ли не единственно реальной основой для упрочения социальных связей были стихийно возникшие узы естественного кровного
f, родства. Во-вторых, можно думать, что при неупорядоченности полового общения, и, следовательно, отсутствии понятия отцовства отношения родства должны были устанавливаться только между потомками одной матери, - Т. е. строиться по материнской, женской линии. Наконец, в-третьих, наиболее стабильной частью тогдашних коллективов были женщины, игравшие важную роль во всех / областях хозяйственной жизни и исключительную роль в
'*• 239
 заботе о детях, поддержании огня, ведении домашнего хозяйства. В силу этих обстоятельств первой упорядоченной формой организации общества, непосредственно сменившей первобытное человеческое стадо, вероятно, был коллектив родственников, связанный общим происхождением по материнской линии, т. е. материнский род.
заботе о детях, поддержании огня, ведении домашнего хозяйства. В силу этих обстоятельств первой упорядоченной формой организации общества, непосредственно сменившей первобытное человеческое стадо, вероятно, был коллектив родственников, связанный общим происхождением по материнской линии, т. е. материнский род.
)
Брак и семья
Многие исследователи считают, что первой исторической формой общественного регулирования отношений между полами был экзогамный дуально-родовой… • ная экзогония), в-третьих, требование супружеской общности. Универсальность… К числу таких институтов относятся прежде всего брачные классы австралийцев. Так, у австралийцев Западной Виктории…Общественные отношения
В коллективной собственности находилась прежде всего земля, промысловая территория со всеми имевшимися в ее пределах объектами охоты, рыболовства и… % 245 дуальным особенностям владельца. Но и эти орудия производства обычно использовались в коллективе и всегда для…Духовная культура
Еще сравнительно недавно считалось, что языки наивнее развитых групп человечества обладают очень не-ачительным, едва ли не в несколько сотен слов,… Развитие языка шло параллельно увеличению объе-!'„ма информации и, в свою… * 253Зак. 486
Возникновение других видов искусства прослеживается этнографически. В устном творчестве раньше всего развились предания о происхождении людей и их… Таким образом, в духовной культуре ранней родовой общины с самого начала… t достигли такой ступени развития интеллекта, когда . потребность объяснить все, с чем им нрихо-| дятся сталкиваться,…II
ческие обряды. В целом тотемизм был своеобразным иде1 ологическим отражением связи рода с его естественной средой, связи, осознававшейся в единственно понятной в то время форме кровного родства.
Анимизм (anima — душа) — вера в сверхъестественные существа, Заключенные в какие-либо тела (души) или действующие самостоятельно (духи). Английским этнографом Э. Тэйлором была выдвинута так называемая анимистическая теория происхождения религии, согласно которой вера в нематериальные души и духов была древнейшим видом религиозных представлений, возникшим в сознании «философствующего дикаря» при попытках обьяснитьтакие явления, как сон, обморок, смерть. В действительности подобные верования не могли быть исходным видом религии, так как представления о нематериальном предполагают известное развитие абстрактного мышления. Но какие-то зачатки анимистических верований в форме смутного одушевления природы, несомненно, появились уже в самую раннюю пору родового строя. У тасманийцев, австралийцев, огнеземельцев и других наиболее примитивных народов* имелись неясные представления о душах живых и умерших людей, злых и добрых духах, обычно мыслившихся в качестве физических, осязаемых существ. Можно думать, что с этими представлениями были как-то связаны и ранние формы почитания матерей-охранительниц очага, засвидетельствованные находками позднепалеолитйческих женских статуэток.
Фетишизм (fetico — талисман, амулет) — вера в Сверхъестественные свойства неодушевленных предметов, например, определенных орудий труда, предметов обихода, деревьев, камней, пещер, а позднее и специально изготовленных культовых предметов. Существует мнение, что простейший фетишизм, состоящий в наделении некоторых предметов ближайшего окружения наряду с обычными также и чудодейственными свойствами, мог быть начальной формой религии. Так, фетишем могло стать
) добычливое копье или плодоносящее дерево, насытившее людей после длительной голодовки. Однако проверить справедливость этой теории трудно: хотя элементы фетишизма широко известны среди отсталых племен,
|; они переплетаются здесь с элементами тотемизма, анимизма и других религиозных представлений.
Магия (mageia — колдовство) — вера в способность
I человека особым образом воздействовать на других людей, животных, растения, явления природы. Не понимая
Г настоящей взаимозависимости наблюдаемых фактов и явлений, превратно истолковывая случайные совпадения, первобытный человек полагал, что с помощью определенных приемов — действий и слов — можно вызвать дождь
| или поднять ветер, обеспечить успех в охоте или собирательстве, помогать или вредить людям. Большое распространение получила, в частности, производственная, или промысловая, магия: такова, например, «пляска кенгуру» у австралийцев, во время которой одни исполнители изображали этих животных, а другие якобы поражали их ко-
|' пьями. По мнению некоторых ученых, практика промысловой магии археологически засвидетельствована частыми знаками ран на позднепалеолитйческих рисунках и скульптурах животных. Рано развились и другие основные виды магии: вредоносная — наведение «порчи» на врага, охранительная —предотвращение этой порчи, лечебная — колдовское лечение ран и недугов.
Таким образом, уже самые ранние виды религии заключали в себе начатки не только фантастических представлений — веры, но и священнодействий ■— культовой практики. Последняя долгое время не составляла тайны: совершение религиозных церемоний было доступно всем и каждому. Но с развитием верований и усложнением культа его отправление потребовало определенных знаний, умений, навыков, опыта. Важнейшие культовые действия стали совершаться старейшинами (например, у сеноев), а затем и особыми специалистами (у огнеземель-
<, 261




 цев, семангов и др.). Как показывают мифы австралийцев и обычаи ряда других племен, в качестве таких специалистов первоначально выступали преимущественно женщины.
цев, семангов и др.). Как показывают мифы австралийцев и обычаи ряда других племен, в качестве таких специалистов первоначально выступали преимущественно женщины.
Религия играла очень большую роль в жизни родовой общины. Видный французский этнограф Л. Леви-Брюль, изучая духовную культуру отсталых племен, даже пришел к выводу, что мышление первобытных коллективов было всецело иррациональным, мистическим, религиозным. Это, конечно, неверно, так как если бы древнейшее человечество руководствовалось только превратными представлениями, оно остановилось бы в своем развитии или погибло. В действительности духовной культуре ранней родовой общины было присуще тесное переплетение рациональных и превратных представлений. Так, леча рану, первобытный человек обычно обращался и к полезным травам и к маши; протыкая копьем изображение животного, он одновременно практиковался в приемах охоты или показывал их молодежи и магически обеспечивал успех предстоящего дела. Это переплетение нередко служит основой для различных построений, выводящих из религии другие явления духовной культуры и идеологии, в особенности искусство и нравственность. Но различные формы общественного сознания, хотя они всегда находились в активном взаимодействии, не могли возникать одна из другой, так как их общей основой были условия общественного бытия.
Таким образом, первобытнообщинная формация была самой длительной в истории человечества. Ее нижняя граница по новейшим данным — примерно 2 млн лет назад, верхняя граница колеблется в пределах последних 5 тыс. лет; в Азии и Африке первые цивилизация возникли на рубеже 4—3 тысячелетий до н. э.; в Европе — в 1 тысячелетии н.э., в других областях Земли — еще позднее.




 Этническая антропология, или расоведение, изучает антропологический состав народов земного шара в настоящем и прошлом. Полученные в результате этого изучения материалы дают возможность выяснить родственные отношения между расами и историю их возникновения, т. е. древность, место и причины происхождения расовых типов.
Этническая антропология, или расоведение, изучает антропологический состав народов земного шара в настоящем и прошлом. Полученные в результате этого изучения материалы дают возможность выяснить родственные отношения между расами и историю их возникновения, т. е. древность, место и причины происхождения расовых типов.
Этническая антропология обслуживает также интересы и смежных с ней дисциплин, разрешая при этом ряд различных задач:
1. Сравнительное изучение человеческих рас весьма важ
но для разрешения проблемы происхождения человека
современного вида и в первую очередь для разрешения
вопроса о том, в одном или нескольких местах возник
человеческий род. В истории науки этот вопрос всегда
рассматривался и решался в неразрывной связи с воп
росом о степени взаимного сходства и различия между
человеческими расами, а также об изменчивости расо
вых типов под влиянием географической среды.
2. Этническая антропология представляет существенный
материал для изучения закономерностей роста, измен
чивости и корреляций, весьма важных для биолога.
3. Антропологический (расовый) материал имеет боль
шое значение и для разрешения некоторых вопросов
истории, в особенности этногенеза. Наличие на тер
ритории типа, резко отличного от основной массы на
селения, позволяет с уверенностью говорить о при
ходе на данную территорию либо предков основной
массы населения, либо предков типа, составляющего
меньшинство. Более того, в этнографических и архе
ологических данных в подобных случаях всегда об
наруживаются различия в культуре в настоящем или
в прошлом между группам^ отличающимися друг
от друга по антропологическому типу. Совокупными
усилиями антрополога, этнографа, археолога, а иног
да и лингвиста можно определить область, из кото
рой шло переселение, и время этого переселения.
Глава 9. ОБЩЕЕ ПОНЯТИЕ О PAGE
 Все ныне живущее человечество представляет собой с биологической точки зрения один вид Homo sapiens. Этот вид распадается на целый ряд более мелких делений, называемых расами. Представителями наиболее резко различающихся расовых типов могут служить, например, суданский негр, монгол, прибалтийский европеец. Основ ной особенностью расовых вариаций, в отличие от всех других внутривидовых вариаций, является географический ареал их распространения. В пределах этого ареала сосредоточена обычно большая часть представителей данного расового типа. В зоогеографии под ареалом распространения подразумевают определенную территорию на земной поверхности, которая получается, если обвести на карте крайние точки, где встречается данное животное.
Все ныне живущее человечество представляет собой с биологической точки зрения один вид Homo sapiens. Этот вид распадается на целый ряд более мелких делений, называемых расами. Представителями наиболее резко различающихся расовых типов могут служить, например, суданский негр, монгол, прибалтийский европеец. Основ ной особенностью расовых вариаций, в отличие от всех других внутривидовых вариаций, является географический ареал их распространения. В пределах этого ареала сосредоточена обычно большая часть представителей данного расового типа. В зоогеографии под ареалом распространения подразумевают определенную территорию на земной поверхности, которая получается, если обвести на карте крайние точки, где встречается данное животное.
Определение того, чем является человеческая раса, следует строить на основе анализа процесса расообразова-ния у человека. Этот процесс протекал под влиянием двух различных факторов: биологического и социального. Роль того и другого становится ясной, если различать в процессе расообразования: 1) механизмы этого процесса и 2) конкретную историю их проявления во времени и пространстве. Механизмы образования каждого отдельного расового признака относятся у человека к области биологии. Наоборот, конкретная история осуществления всех этих механизмов сочетания отдельных признаков в комплексы, или расовые типы, относится у человека к области его
% 265



 социальной жизни. Так, например, для понимания механизма расщепления признаков человека, определяющих цвет и форму его волос, достаточно знания некоторых общих и частных положений генетики. Для понимания причин проникновения, например, тугой формы волос за пределы Восточной Азии на о-в Мадагаскар, необходимо знать историю заселения этих областей, а следовательно, те социальные причины, которые его вызвали.
социальной жизни. Так, например, для понимания механизма расщепления признаков человека, определяющих цвет и форму его волос, достаточно знания некоторых общих и частных положений генетики. Для понимания причин проникновения, например, тугой формы волос за пределы Восточной Азии на о-в Мадагаскар, необходимо знать историю заселения этих областей, а следовательно, те социальные причины, которые его вызвали.
Расы животных и человека
подвида, а каспийский — ни одного. Белый хорек имеет много подвидов, а черный — ни одного. Расы млекопитающих отличаются обычно раскраской шерсти или кожи, размерами и… Во многих случаях отнесение той или иной группы к категории вида или расы представляет трудности, понятные с точки…Раса и нация
Необходимо строго различать два весьма разных понятия — нация и раса. Людей объединяют в нации общность языка, территории, экономической жизни,… Отсутствие внутренней необходимой связи между расой и языком вытекает с… v, 269Расообразование
1) возникновение признаков; 2) закрепление признаков и сохранение их в населении; 3) распространение признаков на территории;Возникновение признаков
косвенные методы, которые позволили по отношению к человеку вычислить, например, приблизительную частоту появления мутаций, определяющих гемофилию.…Закрепление признаков
V 273 не обладают. В человеческой эволюции естественный отбор, очевидно, не мог… верхнего палеолита особенности кожи. Возможно, что более темная пигментация кожи негрской, эфиопской, индонезийской,…V 277


 одни расы ныне характеризуются большими ареалами, я другие — малыми, то источники этих различий лежат не в особенностях самих рас, а в различиях демографических показателей типов хозяйства (охотничьего, земледельческого, скотоводческого и др.) и, до известной степени, в географических особенностях территории, определяющих возможность ее использования.
одни расы ныне характеризуются большими ареалами, я другие — малыми, то источники этих различий лежат не в особенностях самих рас, а в различиях демографических показателей типов хозяйства (охотничьего, земледельческого, скотоводческого и др.) и, до известной степени, в географических особенностях территории, определяющих возможность ее использования.
Смешение рас
Метисная популяция по средней арифметической величине, вычисленной для каждого измерительного признака и для признака, определяемого баллом,… Встречаясь с какой-либо группой, история формирования которой не выяснена,… Так, например, различные народы Западной Сибири обладают в общем менее резко выраженными чертами мон-Глава ТО. КЛАССИФИКАЦИЯ НАРОДОВ МИРА
 10.1. Антропологическая классификация
10.1. Антропологическая классификация
Антропология исследует физические различия между людьми, исторически сложившиеся в ходе их развития в различной естественно-географической среде. Физические различия между группами людей, живущими на значительном друг от друга расстоянии, в разных природных условиях, называются расовыми. Расы постоянно смешиваются, поэтому они имеют не столько наследственный, сколько исторически сложившийся характер. Антропология считает расы совокупностью территориальных групп людей, а не отдельных особей. По всем главным морфологическим, физиологическим и психологическим особенностям, которые характеризуют современных людей, сходство между расами очень велико, а различия не существен-
| вы. Можно полагать, что современные расы начали складываться в позднем палеолите, и с тех пор они все время изменяются вследствие смешений и других причин. Расовые признаки не имели и не имеют никакого значения
|'" для исторического развития людей, уровня развития разных групп человечества, их культуры.
Признаки, по которым выделяются расы, очень различны. Одни из них описательные, другие сугубо специ-1 альные. Существенным расовым признаком является фор-
V 281


 ма волос головы и степень развития третичного волосяного покрова (усы, борода). У большей части обитателей Европы волосы обычно мягкие, прямые или волнистые. У монголоидов, обитателей Восточной и Юго-Восточной Азии и ряда других областей мира они тугие и прямые. У африканцев — жесткие и курчавые. Третичный волосяной покров сильно развит у обитателей Европы, Передней Азии, Закавказья, коренного населения Австралии. Значительно слабее он выражен у многих групп африканцев, населения некоторых частей Азии.
ма волос головы и степень развития третичного волосяного покрова (усы, борода). У большей части обитателей Европы волосы обычно мягкие, прямые или волнистые. У монголоидов, обитателей Восточной и Юго-Восточной Азии и ряда других областей мира они тугие и прямые. У африканцев — жесткие и курчавые. Третичный волосяной покров сильно развит у обитателей Европы, Передней Азии, Закавказья, коренного населения Австралии. Значительно слабее он выражен у многих групп африканцев, населения некоторых частей Азии.
Определенное значение при расовой оценке имеет окраска (пигментация) кожи и волос. Так, слабо пигментированы некоторые группы населения Северной Европы. Темная окраска кожи и волос свойственна большей части африканцев. Темные волосы, но более светлая пигментация кожи у обитателей Южной Европы, у значительной части коренного населения Азии.
Несколько меньшее значение как расовому признаку придается росту, весу и пропорциям тела. По росту и пропорциям тела выделяются главным образом некоторые малорослые (пигмейские) группы в Южной Африке и негритосы Юго-Восточной и Южной Азии.
Немалое значение для выделения расовых типов имеет изучение признаков головы и лица. Но головной указатель, т. е. соотношение длины и ширины черепа, очень сильно варьируется в каждой расе и сам по себе расовым показателем не является. Имеют классификационное значение и некоторые другие признаки, например, форма носа.
Для антропологических исследований привлекаются и многие специальные признаки: данные о генах, группах крови, узорах на пальцах (папиллярные узоры), форме зубов и др.
Согласно классификацииЯ.Я. Рогинского выделяют три большие расы: австрало-негроидную, или экваториальную, евразийскую, или европеоидную, и азиатско-аме-
' риканскую, или монголоидную. Промежуточное место между экваториальной и евразийской большими расами занимают две расы: эфиопская и южноиндийская; между евразийской и азиатско-американской размешаются также две расы — южносибирская и уральская; особое место по значительной части признаков, промежуточное между американской и австралийской, занимают полинезийская и курильская расы.
Экваториальная большая раса
1. Австралийская раса характеризуется темной кожей, волнистыми волосами, обильным развитием третич ного волосяного покрова на лице и на теле, очень… 2. Ведцоидная раса отличается от предыдущей слабым развитием третичного… V 283S4
3. Меланезийская раса (включая негритосские типы), в
отличие от двух предыдущих, обладает курчавыми
волосами; по обильному развитию третичного воло
сяного покрова, по сильно выступающим надбровным
дугам некоторые ее варианты весьма сходны с авст
ралийской расой. По своему составу меланезийская
раса гораздо более пестрая, чем негрская.
4. Негрская раса отличается от австралийской и веддо-
идной (и в гораздо меньшей степени от меланезийс
кой) очень сильно выраженной курчавостью волос;
от меланезийской она отличается большой толщиной
губ, более низким переносьем и более плоской спин
кой носа, несколько более высокими орбитами, мало
выступающими дугами и, в общем, более высоким
■ ростом.
5. Негрилльская (центральноафриканская) раса отлича
ется от негрской не только очень низким ростом, но
также более обильным развитием третичного воло
сяного покрова, более тонкими губами, более резко
выступающим носом и более высоким носовым по
казателем. _ ■ '
6. Бушменская (южноафриканская) раса отличается от
негрской не только очень низким ростом, но и более
светлой кожей, более узким носом, более плоским
лицом, весьма уплощенным переносьем, малыми
размерами лица и, кроме того, стеатопигией (отло
жением жира в ягодичной области) и другими осо
бенностями. Иногда встречается эпикантус.
Евразийская большая раса
Евразийская, или европеоидная, большая раса характеризуется вообще светлой или смуглой окраской кожи, прямыми или волнистыми мягкими волосами, обильным ростом бороды и усов, узким, резко выступающим носом, высоким переносьем, сагиттальным расположением
V 285
 | |||
 | |||

|

|
э, тонкими губами. Из краниологических признаков для нее характерны: малый или средний н^осовой указатель, высокий ринальный указатель, высокий симотическнй указатель, ортогнатизм, малый угон горизонтальной профилировки, среднее или сильное развитие кяыковой ямки. Для северо-западных вариантов это^ расы типичны евет лью глаза и волосы.
Область распространения —. Европа, Северная Африка, Передняя Азия, Северная Ин^дия.
Европеоидная раса подразделяется на ряд малых рас (рис, 46).
1. Атланто-балтийская раса характеризуется светлой ко
жей, светлыми волосами ^ глазами, большой дли
ной носа (в процентах нижнего отдела лица), высо-
ким ростом.
2. Среднеевропейская раса «Обладает в общем менее
светлой пигментацией волосу и глаз, чем предыдущая,
несколько меньшим pocroKi.
3. Индо-средиземноморская jjjaca характеризуется тем
ной окраской волос и глаз,, смуглой кожей, волнис
тыми волосами, еще более 5 удлиненным носом, чем
нредыдущие, несколько б^одее выпуклой спинкой
носа, очень узким лицом, д^олихо- и мезокефалией.
4. Балкано-кавказская раса харчэакгеризуетсятемньши во
лосами, темными или смешанными глазами, выпук-
лым носом, очень обильном развитием третичного
волосяного покрова, сравнительно коротким и очень
широким лицом, брахикефоалией, высокимростом.
5. Беломорско-балтийскаярас^а— очень светлая, однако
несколько более пигментир^ованная, чем атланто-бал-
тийская, обладает средним ; (а не сильным) ростом бо
роды, имеет сравнительно короткий нос с прямой или
вогнутой спинкой и значиге^льнымлроцентом припод
нятых оснований, короткое^ лицо и средний рост.



Рве. 46. Евразийская большая раса:
1 — атланто-балтийская; 2 — среднеевропейская;3'—нндо-средиземноморская; 4—балкано-кавказская; 5 — беломор-ско-балтийская
V 287

|

|
Азиатско-американекая раса
Ареал азиатско-американской расы охватывает Восточную Азию, Индонезию, Центральную Азию, Сибирь, Америку. Азиатско-американская раса подразделяется на несколько малых рас (рис. 47). … 1. Североазиатская раса выделяется среди азиатско-аме-риканских рас преимущественно меньшим процентом тугих волос,…Зак. 486
Центральноазиатский тип представлен в различных вариантах, из которых некоторые сближаются с байкальским типом, другие — с вариантами арктической и… 2. Арктическая (эскимосская) раса отличается от севе роазиатской более… 3. Дальневосточная раса по сравнению с североазиат ской также характеризуется более тугими волосами, и темной…Географическая классификация
Географическая классификация предусматривает распределение человечества по отдельным группам и народам в соответствии с местами их обитания. Но при этом она не учитывает их происхождения, уровня развития хозяйственных и социальных отношений, облика культуры. Поэтому географическая классификаций больше используется при описании народов, чем при их исследовании. Вместе с тем расселенные по соседству народы нередко имеют обшее происхождение и сходную культуру.
Австралия и Океания
лее изучены: курнаи и нарриньери — на юго-востоке Австралии, камиларои на востоке, племена аранда (арунта), диэри, лоритья, арабана (урабунна) в… В антропологическом отношении большинство аборигенов Австралии относится к… Тасмания. Тасманийцы вымерли в конце прошлого столетия в результате интенсивной колонизации европейцами. Последняя…Азия
Индонезия, или Малайский архипелаг, географически включает Большие Зондские о-ва — Суматра, Ява, Калимантан (Борнео), Сулавеси (Целебес); Малые Зондские о-ва — Бали, Сумбава, Флорес, Тимор и др.; Молуккские о-ва и ряд других мелких островных групп. Численность населения Индонезии превышает 155 млн человек.
В языковом отношении население Индонезии однородно и говорит на индонезийских языках, принадлежащих к обширной австралонезийской (малайско-полинезин-ской) языковой группе.
Две трети населения Индонезии живет на о-ве Ява, где плотность его достигает очень больших размеров (до 1 500 человек на км2). Наиболее многочисленная группа яванцы (свыше 62 млн), заселяющие центральную часть острова; к ним близки сунды, живущие в западной части Явы (18,5 млн), и мадурцы на о-ве Мадура и восточной Яве (9 млн). В горных районах островов сохранились малочисленные племена тештеров и бадуев.
Население Суматры, приближенных районов Калимантана, Сулавеси и других островов известно в литературе под названием малайцев. Малайский язык широко распространен во всей Индонезии и признан государственным языком.
Остров Суматра наряду с Явой — область высокой древней культуры и центр, откуда шло расселение малайцев на другие острова. Наиболее крупными группами на Суматре являются малайцы и близкие к ним минангкабау и ред-жанг-лампунги; на северном побережье острова расселены аче (2,1 млн). Более низок уровень культуры у батаков (3,1 млн). В неблагоприятных болотистых районах восточной Суматры сохранились одни из наиболее отеталых~в Индонезии племен — кубу, лубу, улу и др. — охотники и собиратели; их общая численность около 20 тыс. человек.
Прибрежные районы Калимантана заселены пришельцами с других о-вов — Явы, Суматры, Сулавеси. В ос-
v 299

 новном это малайцы, бути и яванцы. Древними жителями острова являются: нгаджу, кайяны, кенья, бахау, ке-даяны, муруты, дусуны — племена мотыжных земледельцев, охотников и рыболовов, объединяемые под общим Названием «даяки».
новном это малайцы, бути и яванцы. Древними жителями острова являются: нгаджу, кайяны, кенья, бахау, ке-даяны, муруты, дусуны — племена мотыжных земледельцев, охотников и рыболовов, объединяемые под общим Названием «даяки».
Население о-ва Сулавеси представлено несколькими группами. В южной части острова расселены бугй и мака-сары (5 млн человек), на севере — минахасы-горонтальцы (1 млн). Племена центральных областей сулавеси известны под общим названием «тораджи» (1,4 млн).
Коренные обитатели Андаманских о-вов — андаман-цы — в настоящее время почти полностью уничтожены европейской колонизацией, хотя в середине XIX в. они составляли единственное население этих островов, и их численность достигала 5 тыс. человек. Язык андаманцев занимает изолированное положение среди языков Южной Азии.
Население Филиппинских о-вов, вытянутых с юга на север более чем на 1 800 км, насчитывает свыше 44 млн человек. Оно говорит на языках индонезийской группы, близких к языкам Малайского архипелага. Из наиболее крупных групп филиппинцев можно назвать висайя (20 млн); тагалов — на о. Лусон, штоков. Эти народы населяют прибрежные равнинные районы и имеют развитую земледельческую культуру. Во внутренних горных областях живут: ифугао, илонготы — на Лусоне; манги-ан, тагбануа—в центральных районах о. Мандоро.
Особую Fpynny, резко отличную по своему антропологическому типу, составляют аэта — охотники и собиратели, сохранившиеся преимущественно в некоторых районах Лусона, Бисайских о-вов, Минданао, Палавана.
В населении Малайского архипелага можно различить, если отвлечься от более поздних примесей (западноевропейской, китайской, индийской и арабской), три основных антропологических «пласта»: наиболее поздний представлен южноазиатской расой (малайцы, яванцы, сун-
ды); предшествующий пласт относится к той же южноазиатской расе, но в подавляющей массе — к ее более низкорослому, длинноголовому и широконосому варианту, характеризующемуся также более коротким лицом и меньшим процентом эпикантуса (ифугао, тогалы, даяки). Еще более ранний пласт составляют волнистоволосые, темнокожие типы, относящиеся к ведцоидной расе (тоалы и лой-нанги Сулавеси, орангбатины восточной Суматры, малац-ги, кайаны, улуайары Калимантана).
К тому раннему пласту относятся также весьма низкорослые варианты меланезийской расы среди аэта и андаманцев. Изученные группы аэта, по-видимому, сложны по составу. Так, у мужчин аэта из восточной части Лусона наряду с курчавыми волосами примерно в 18% встречаются волнистые волосы; иногда у них имеется значительный рост бороды, губы обычно средней толщины, носовой указатель невелик — 79, эпикантус отсутствует.
Вопрос о том, какая из двух рас: веддоидная или меланезийская (в ее низкорослом варианте) — является более древней на о-вах Индонезии, решается в антропологии неодинаково. Совокупность имеющихся фактов свидетельствует, скорее, о древности темнокожего, волнис-товолосого, а не курчавого населения. Возможно предполагать, что меланезийская раса возникла как местный вариант австралийской расы. Однако при малой еще изученности древнейшей истории негрских народов Африки нельзя считать вполне исключенной гипотезу и о древних миграциях в Меланезию из Восточной Африки.
Индокитай. Народы Индокитая относятся к 4 основным языковым семьям: австроазийской (мон-кхмерская и вьетнамская группы и группа мяо-яо), таи, австронезийской и китайско-тибетской. Народы мон-кхмерской группы принадлежат к древнему населению Индокитая; еще в I тысячелетии они занимали большую часть полуострова и образовывали здесь крупные государства. В дальнейшем народы мон-кхмерской группы были оттеснены
V 301





 и в значительной степени ассимилированы продвигающимися с севера народами китайско-тибетской группы.
и в значительной степени ассимилированы продвигающимися с севера народами китайско-тибетской группы.
В настоящее время к мон-кхмерским народам относятся кхмеры в Камбодже, Южном Вьетнаме и Южном Лаосе, моны — в Южной Бирме; ва и палаунг — в северных частях Бирмы и Таиланда и различные племена (сти-енги, мнонги, чамре, банар, кун, сун и др.), объединяемые под общими названиями горных кхмеров и монов в горных областях Вьетнама, Камбоджи и Лаоса. В языке и культуре этих племен отмечаются черты сходства с индонезийцами.
Самый многочисленный народ Индокитая —вьетнамцы (вьеты)—основное население Вьетнама. Вьетнамский язык обнаруживает черты сходства с языками мон-кхмер-ской группы и языками индонезийскими. Вьетнамцы сложились в результате ассимиляции древнего населения Западного Индокитая — мон-кхмерских и индонезийских групп — племенами, распространившимися из Юго-Восточного Китая.
К группе тан относятся сиамцы — основное население Таиланда, лао — в Лаосе, шань — в Северо-Восточной Бирме, карены—в Восточной Бирме и Западном Таиланде. По своему происхождению народы группы тан связаны с Южным Китаем.
В антропологическом отношении население п-ва Индокитай относится к двум большим расам: монголоидной и австрало-негроидной. Территория Индокитая издавна являлась областью их контакта и смешения. Численно резко преобладают монголоидные типы, которые характерны для вьетнамцев, сиамцев, бирманцев и многих других групп. Эти типы принадлежат к южноазиатской расе, в пределах которой могут быть выделены различные варианты.
На п-ве Малакка австрало-негроидная большая раса представлена в двух вариантах: веддоидной и меланезийской расами. Первый характерен для сеноев, второй — для семангов. Основное отличие семангов от типичных пред-
ставителей меланезийской расы — малый рост (152 см) и связанные с ним другие особенности (низкое лицо, слабые надбровные дуги); у семангов отмечается менее обильный третичный нокров.
Черты веддоидной расы в виде примеси проявляют-| ся в населений Индокитая и далее к северу. Так, у некоторых племен мои встречаются волнистые волосы, сравнительно толстые губы, широкие носы и более темная кожа.
Китай. По своему антропологическому тину китайцы относятся к дальневосточной расе: черты ее в наиболее выраженном виде выступают у населения Северного Китая. Для северных китайцев характерны: темная пигментация, прямые жесткие волосы, слабое развитие третичного волосяного покрова, хорошо выраженные монголоидные особенности глаз, высокое и сравнительно узкое лицо, сравнительно узкий нос, умеренно толстые губы,
Южные китайцы отличаются от северных более темной пигментацией, большим процентом волнистых волос, несколько более сильным развитием третичного волосяного покрова, менее монголоидным строением глазной области, более широким носом, более толстыми губами, более прогнатным и менее высоким лицом, более низким ростом. Такое направление изменчивости свидетельствует о наличии признаков южноазиатской расы и отражает этническую историю Южного Китая.
Помимо основного китайского населения на территории Китая проживает ряд других народностей и этнических групп. Большая часть некитайского населения по количеству народностей и племен приходится на центрально-южные и юго-западные районы страны. Основная масса их относится к трем лингвистическим группам: мяо-яо, тибето-бирманской и таи.
В первую группу входят народы мяо и яо (в провинции Гуйчжоу и др.) и некоторые другие народы, у которых преобладают черты южноазиатской расы.
v 303

 К тибето-бирманской группе относятся и, или, ицзу, наси, бай, туцзя, лису и другие, живущие преимущественно в провинциях Юньнань, Сычуань, на северо-западе Гу-анси и юго-востоке Хунани. Следует особо вьщелить антропологические особенности ицзу. В антропологической литературе распространено мнение о европеоидных особенностях этих племен: при этом ссылаются на такие признаки, как выступающий нос, широкий разрез глаз, сравнительно светлая кожа. Мнение это не является достаточно убедительным: указанные признаки не могут служить доказательством европеоидной примеси среди ицзу, ко-1 торые по своему антропологическому типу близки, вероятно, к седангам ибаанарам п-ва Индокитай (Рогинский, Левин, 1978). У ицзу сочетаются очень тугие волосы со слабым развитием бороды и малым процентом эпиканту-са и довольно резко очерченным носом.
К тибето-бирманской группе относятся и, или, ицзу, наси, бай, туцзя, лису и другие, живущие преимущественно в провинциях Юньнань, Сычуань, на северо-западе Гу-анси и юго-востоке Хунани. Следует особо вьщелить антропологические особенности ицзу. В антропологической литературе распространено мнение о европеоидных особенностях этих племен: при этом ссылаются на такие признаки, как выступающий нос, широкий разрез глаз, сравнительно светлая кожа. Мнение это не является достаточно убедительным: указанные признаки не могут служить доказательством европеоидной примеси среди ицзу, ко-1 торые по своему антропологическому типу близки, вероятно, к седангам ибаанарам п-ва Индокитай (Рогинский, Левин, 1978). У ицзу сочетаются очень тугие волосы со слабым развитием бороды и малым процентом эпиканту-са и довольно резко очерченным носом.
В Северо-Западном Китае живут уйгуры, киргизы, казахи, говорящие на языках тюркской группы, и хуэй, по языку и культуре близкие к китайцам, но принявшие ислам. У уйгуров Синьцзяна преобладает темнопигменти-рованный брахикефальный европеоидный тип, близкий, по-видимому, к типу среднеазиатского междуречья, характерному для ряда народов Средней Азии.
Значительное большинство населения Тибета — тибетцы (бодо), говорящие на языке тибето-бирманской группы. Тибетцы по своему антропологическому типу суммарно близки к северным китайцам. Среди них отмечают два варианта: один, характеризующийся более выступающим носом, более массивный, с меньшей высотой черепа и с несколько ослабленными монголоидными чертами, локализован на востоке, другой обнаруживает особенности южноазиатской расы и распространен на юге страны.
Монголия.Более 90% населения страны составляют монголы (в основном халха-монголы, дербеты, торгоу-ты). По своему антропологическому типу монголы относятся к центрально-азиатскому варианту североазиатской
расы. Можно предполагать наличие здесь примеси даль-£ невосточной расы.
Корея.Население Кореи по своему этническому со
ставу однородно. Корейский язык занимает изолирован
ное положение в лингвистической классификации; неко
торые авторьГуказывают на его связь с тунгусо-маньчжур
скими языками. >
По антропологическому типу корейцы относятся к дальневосточной расе. Обнаруживая большое сходство с северными китайцами, корейцы отличаются от них поми-| мо брахикефалии рядом особенностей. Такие признаки, отличающие корейцев от северных китайцев, как усиление волосяного покрова, увеличение ширины носа, увеличение толщины губ и некоторые другие, свидетельствуют о наличии в антропологическом типе корейцев элементов южноазиатской расы, обнаруживающихся также в типе южных китайцев. С другой стороны, для корейцев характерны крупные абсолютные размеры лица и некоторая депигментация, что указывает на связи с североазиатскими монголоидами.
Япония.Этнический состав населения Японии однороден. Положение японского языка в лингвистической классификации пока еще не ясно. Как в языке, так и в культуре японцев помимо сильного китайского влияния отмечаются элементы южного происхождения.
Антропологический тип японцев в целом может быть охарактеризован как смешанный. От типичных представителей дальневосточной расы — китайцев и корейцев — японцы отличаются рядом признаков, одни из которых сближают их с айнами (большее развитие волосяного покрова, значительный процент волнистых волос), другие с айнами июжными монголоидами (более темная кожа, широкий нос, толстые губы, низкое лицо, малый рост — 158—160 см). Эти данные свидетельствуют об участии в формировании антропологического типа японцев помимо дальневосточной расы также южноазиатской и курильской.
V 305





 чает казахов, киргизов, узбеков, таджиков, туркмен, уйгуров, каракалпаков. По своему антропологическому типу казахи и киргизы в целом сходны между собой и относятся к южносибирской расе, занимающей по основным ра-сово-диагностическим признакам промежуточное место между азиатско-американской и евразийской большими расами.
чает казахов, киргизов, узбеков, таджиков, туркмен, уйгуров, каракалпаков. По своему антропологическому типу казахи и киргизы в целом сходны между собой и относятся к южносибирской расе, занимающей по основным ра-сово-диагностическим признакам промежуточное место между азиатско-американской и евразийской большими расами.
Казахи характеризуются значительно повышенным по сравнению с монголоидами ростом бороды, сравнительно выступающим носом с высоким переносьем, ослабленным развитием эпикантуса; волосы у них прямые, умеренно жесткие; пигментация умеренно темная (до 50% смешанных и светлых оттенков глаз); лицо высокое, очень широкое, уплощенное; голова брахикефальная (головной указатель — 84—85). При очень широком расселении казахи разных районов не обнаруживают существенных различий в своем антропологическом типе.
Киргизы, по сравнению с казахами, характеризуются значительно большим развитием эпикантуса и несколько более выраженной монголоидностью типа в целом. Следует отметить некоторое усиление европеоидных черт у киргизов Ферганской долины по сравнению с киргизами Тянь-Шаня: у первых лицо значительно уже (144,1 мм), развитие бороды несколько больше, процент эпикантуса меньше, лереносье выше.
Уйгуры, живущие на территории Алма-Атинской области (основная их масса живет в Китае), отличаются от соседних с ними казахов более европеоидными чертами; лицо у них ниже и много уже, борода растет сильнее; они гипербрахиальны (головной указатель — 87).
Таджики являются представителями антропологического типа, описанного в литературе под названием «пами-ро-ферганского». Он в известной степени сближается с индо-средиземноморской расой. Для него характерны следующие признаки: темная пигментация, широковол-
ннстые волосы, сильное развитие третичного волосяного покрова, средневысокое лицо, выступающий узкий прямой нос, прямой лоб со слабо развитым надбровьем. Отличительной особенностью этого типа является довольно широкое лицо (135—142 мм), брахикефалия (головной указатель — 83—84). Указанная характеристика в наибольшей степени относится к типу горных таджиков. Равнинные таджики отличаются некоторой примесью монголоидных рас.
Узбеки, особенно узбеки Ферганы, близки ж равнин-ным таджикам; среди них также преобладает тип Среднеазиатского междуречья, но монголоидные черты выражены у узбеков в целом несколько сильнее; у отдельных групп, в особенности у сохранивших родо-племенное деление (кыпчак, мангыт, курама) монголоидная примесь очень значительна; так, у узбеков Хорезма (группа мангыт) 35 % эпикантуса, низкое переносье, ослабленный рост бороды.
Среди туркменов преобладает европеоидный тип, характеризующийся темной пигментацией, сильно выступающим узким носом, сравнительно узким и невысоким лицом, обильным волрсян*ым покровом. Особенностью, отличающей туркменов от представителей памиро-ферган-ского типа, является их долихокефалия, головной указатель — 75—77. Антропологический тип туркменов большинство исследователей сближают с длинноголовым европеоидным типом средиземья, т. е. с индо-средиземноморской расой. У туркменов имеется и монголоидная примесь, выраженная у различных групп в разной степени. Так, туркмены-чаудыры Хорезма более монголоидны по сравнению с туркменами-иомудами (у них 24% эпикантуса, ниже переносье, слабее рост бороды, выше и шире лицо).
Южная Азия.В состав территории Южной Азии входят Индия, Бангладеш, Пакистан. По языку население Индии принадлежит к следующим лингвистическим груп-
 пам: индоарийской (индийской), иранской, дравидийской, мунда и тибето-бирманской. Огромное большинство населения говорит на индоарийских и дравидийских языках.
пам: индоарийской (индийской), иранской, дравидийской, мунда и тибето-бирманской. Огромное большинство населения говорит на индоарийских и дравидийских языках.
Народы, говорящие на индоарийских языках, составляют большинство населения Северной Индии, Бангладеш и Пакистана; народы дравидийской группы расселены в Южной Индии. Народы мунда живут преимущественно в восточной части Центральной Индии, народы тибето-бирманской группы — в областях, пограничных с Китаем и Бирмой. На иранских языках говорят белуджи и различные афганские племена (пуштуны) в соседних с Афганистаном районах.
В антропологическом отношении население Индии неоднородно. Север Индии — область расселения индо-средиземноморской расы, Центральная и Восточная Индия — веддоидной расы, Южная Индия — южноиндийской (дравидийской) расы.
Строгого соответствия между языковыми и антропологическими группами не имеется. Хотя огромную массу индоевропейского населения составляют различные варианты индо-средиземноморскойрасы, однако это справедливо главным образом для Северо-Западной Индии (рад-жастханцы, сикхи, пенджабцы). Для них характерны высокий рост (172 см), долихокефалия, прямая спинка носа. Индоевропейские народы Центральной и Южной Индии (бихарцы, сингалы) обладают ясно выраженной примесью южноиндийской расы; некоторые группы бихарцев ближе всего к веддоидной расе. Аналогичные черты нередки у, индоевропейских народов Западной Индии (маратхи и гуд-жаратцы). Говорящие на языке индоарийской группы бхи-лы (Центральная Индия, Раджастхан) принадлежат в значительной чаоуи к веддоидной расе. Среди дардов и кашмирцев преобладает индо-средиземноморская раса с некоторым процентом светлоглазых и сравнительно светловолосых индивидов.
Дравидийские по языку народы также разнообразны по своему составу. Антропологический тип ораонов и гон-дов имеет много черт, которые сближают его с веддоидной расой. Те же черты имеют дравидийские народности джунглей Центральной Индии. Крупные дравидийские народы Южной Индии (тамилы, телугу) характеризуются признаками южноиндийской расы. По главным признакам эта раса занимает промежуточное положение между индо-средиземноморской и веддоидной, что дает основание считать южноиндийскую расу продуктом смешения двух названных рас.
Передняя Азия— юго-западная часть азиатского материка, включающая Малую Азию, Аравийский п-ов, Иранское нагорье и области, лежащие между ними. Народы Передней Азии относятся по языку в основном к трем группам: семитской, тюркской, иранской. Семитская группа представлена арабскими народами, составляющими основное население Аравийского п-ва и прилегающих стран.
Из народов тюркской группы больше всего в Передней Азии тюрков (анатолийские турки, османы). Турки живут в основном в Турции, где составляют около 85% населения. Проникновение тюркских народов на территорию Малой Азии относится преимущественно к XI в., когда турки-сельджуки — выходцы из Туркмении— покорили местное население.
В антропологическом отношении население Афганистана, значительной части Ирана, части Ирака и большей части Аравийского п-ова относится к различным вариантам индо-средиземноморской расы. Однако в населении областей, примыкающих к Закавказью, ярко выражены черты балкано-кавказской расы, а именно ее переднеази-атского варианта, характеризующегося удлиненным носом, с выпуклой спинкой и опущенным основанием, густыми бровями, очень обильным третичным волосяным покровом, средним ростом (163—167 см). Этот вариант распространен среди армян и ассирийцев. У иранцев Хорасана
v 309
 этот тип отличается удлиненной формой головы. Черты балкано-кавказской расы также выражены у луров и бах-тиаров.
этот тип отличается удлиненной формой головы. Черты балкано-кавказской расы также выражены у луров и бах-тиаров.
Индо-средиземноморская раса характерна для арабов области нижнего течения Евфрата, где она представлена весьма длинноголовым, узколицым, сравнительно высокорослым (168 см) вариантом. Сходный, но несколько менее рослый (164 см) и не столь узколицый тип был описан среда арабов Южной Аравии. Индо-средиземноморская раса хорошо выражена также у многих иранцев и афганцев-патанов (головной указатель — 76, рост —169 см), отличающихся более светлой пигментацией волос и глаз.
Кроме перечисленных антропологических типов в Передней Азии, встречаются смешанные с более далекими формами группы. Так, например, мекраны прибрежных областей Белуджистана, по-видимому, имеют негрскую примесь. Бедуины Гадрамаута обнаруживают черты сходства с веддоидной расой. Хезарейцы Афганистана имеют ясно выраженный монголоидный облик. Эта монголоид-ность, хотя и очень смягченная по сравнению, например, с бурятами, якутами и тунгусами, охватывает целый комплекс признаков: высокий процент прямых волос, очень слабый рост волос на груди и на лице, ушюшенность лица, сильное выступание скул, крупные размеры лица, невысокое переносье, присутствие эпикантуса, малая ширина глазной щели.
Среди турок-османов, обладающих чертами балкансь кавказской расы, имеется примесь депигментированного европеоидного типа и очень легкая монголоидная примесь. Особенно отчетливо видны монголоидные черты у юру-ков.
Кавказ — одна из наиболее сложных по своему этническому составу территорий. Население Кавказа по языку относится к нескольким семьям: 1) собственно кавказских, или иберо-кавказских, языков; 2) индоевропейских языков; 3) тюркских языков.
В основной своей массе, независимо от языковой принадлежности, кавказские народы обладают признаками европеоидной расы. Кроме того, всеони в общем чаше имеют темные волосы, темные или смешанные по цвету глаза, резко выступаннцие носы, часто с выпуклой спинкой и слегка опушенным основанием, сильно развит третичный волосяной покров.
Монголоидные признаки обнаруживаются у народностей степной области Северного Кавказа, в более сильной степени у ногайцев, в более слабой — у туркменов. Ногайцы характеризуются наличием эпикантуса, более низким переносьем, чем, например, у киргизов Тянь-Шаня, слабым ростом бороды, невысоким ростом.
Большая часть населения Кавказа относится к разным типам балкано-кавказской расы. В Восточном Закавказье (Азербайджан) население обладает некоторыми чертами индо-средиземноморской расы. В коренном населении Кавказа В.В. Бунак выделил четыре расы: понтий-скую, каспийскую, понто-загросскую и кавказскую. Первые две включаются им в средиземноморскую расу; две последние — в памиро-альпийскую расу.
Одним из важных обобщений, сделанных грузинскими антропологами, было выделение кавкасионного типа, характеризующегося очень широким лицом (146— 148 мм), относительно светлой пигментацией глаз, высоким ростом (170 см). Он наиболее отчетливо выражен в группах, живущих по склонам Большого Кавказского хребта от андо-цунтинских групп Дагестана на востоке до сванов на западе, включая чеченцев, ингушей, осетин, ка7 рачаевцев и балкарцев. Грузины по раду признаков сходны с народами Северного Кавказа (высокий рост, широкое лицо, не очень темная окраска радужной оболочки).
На Северном Кавказе западные народы имеют значительно меньшие величины головного указателя, чем восточные (например, у адыгейцев — 79, агульцев — 88). В Закавказье наибольшие величины головного показателя
*■ 311

 распространены в центральном нагорье: у пшавов около 86, у армян Карабаха — 86; самые малые величины отмечены в Южном Азербайджане — 76.
распространены в центральном нагорье: у пшавов около 86, у армян Карабаха — 86; самые малые величины отмечены в Южном Азербайджане — 76.
У народов Закавказья скуловая ширина в общем увеличивается с востока на запад: 136 мм у нахичеванских азербайджанцев, 142 — у армян, 143—148 — у грузин. Интенсивность окраски волос и глаз этих народов в общем убывает, а рост увеличивается с востока на запад.
Осетины в общем мало отличаются по типу от народов Северного Кавказа, в частности аварцев*. Значительные отличияот осетин обнаруживает другая иранская группа— курды. По антропологическому типу курды более сходны с тюркоязычными азербайджанцами, чем с ираноязычными осетинами.
Кавказские евреи отличаются средним или ниже среднего ростом, большим головным указателем — от 84 до 88,5; скуловая ширина варьирует от 134 до 144 мм. В общем, евреи Кавказа значительно ближе к брахикефаль-ным нееврейским типам Закавказья, чем к йеменским или польским евреям.
Америка
1) доевропейское население Америки — эскимосы и ин дейцы; 2) потомки европейских иммигрантов, переселивших ся в Америку в разное… 3) потомки невольников, завезенных в Северную и Юж ную Америку из Африки, — негры;V 317



 н плотской группы в Восточном Судане проживают различные кочевые кушитские (бищарин, хадендоа) н арабские племена. В долине Нила, ниже его излучины, расселены нубийцы; язык их относится к северной группе нилотскнх языков. Большая часть населения Восточного Судана говорит на арабском языке, распространение которого связано с проникновением арабов в Восточный Судан.
н плотской группы в Восточном Судане проживают различные кочевые кушитские (бищарин, хадендоа) н арабские племена. В долине Нила, ниже его излучины, расселены нубийцы; язык их относится к северной группе нилотскнх языков. Большая часть населения Восточного Судана говорит на арабском языке, распространение которого связано с проникновением арабов в Восточный Судан.
В отличие от народов Восточного, Центрального и особенно Западного Судана, которые говорят на многочисленных, далеких друг от друга языках, языки народов Центральной и значительной части Южной Африки составляют большую группу языков банту.
По своему антропологическому типу народы банту близки к негрским народам Судана, но в различных областях своего расселения обнаруживают черты смешения с другими антропологическими типами. Так, западные банту, обитающие в тропических лесах бассейна Конго, отличаются от негров Гвинейского побережья более низким ростом, менее выраженной долихокефалией, более обильным третичным волосяным покровом. Имеются основания предполагать смешение с пигмеями — представителями негрилльской расы, для которой характерен очень маленький рост, меньшая долихокефалия, усиленный по сравнению с неграми рост волос на лице и на теле. У отдельных групп восточных банту выявляются черты сходства с народами Северо-Восточной Африки, относящимися к эфиопской расе.
Негрилли, или африканские пигмеи, обладают, как уже указывалось, комплексом признаков, отличающих их от негров, Помимо очень малого роста, более сильного развития третичного волосяного покрова они обладают такими признаками, как очень широкий нос (еще более широкий, чем у негров) с сильно уплощенным переносьем и нередко выпуклой спинкой, тонкие, по сравнению с неграми, губы и несколько более светлая кожа. Смешение
негриллей с их соседями — высокими негрскими группами —привело к значительному разнообразию их антропологического типа.
Бушменская раса составляет основной антропологический элемент у бушменов и готтентотов. Отличительной осо-бенностью их антропологического типа по сравнению с негрской расой являются: более светлая, желтовато-бурая кожа, более тонкие губы, более плоское лицо, наличие в некоторых случаях этжашусаи такие специфические признаки, как морщинистость кожи и стеатопигия.
При общем сходстве антропологического типа бушменов и готтентотов между ними имеются и некоторые различия: у готтентотов несколько более покатый лоб, более сильно развитые надбровные дуги, выше переносье, крупнее размеры черепа, относительно более высокий череп, значительно выше рост. Бушмены, готтентоты и горные дамара относятся к койсанекой языковой группе. В прошлом эти народы занимали почти всю Южную и Восточную Африку; в настоящее время в центральных областях Танзании живут племена хадзапи и сандаве, говорящие на языках, родственных языкам бушменов и готтентотов.
Наиболее типичными представителями эфиопской расы являются народы Эфиопии и п-ова Сомали. Эфиопская раса занимает промежуточное положение между индо-средиземноморской и негрской расами. Примесь эфиопской расы обнаруживается среди некоторых групп восточных банту.
Северная Африка является областью распространения европеоидной большой расы. Естественной границей между европеоидными и негроидными типами является пустыня Сахара. Европеоидные варианты Северной Африки принадлежат к нндо-средиземноморской расе. Сюда относится арабское и берберское население Алжира, Туниса, Марокко, Ливии, кочевые арабские и берберские племена Сахары.
v M9

 Доарабское население Северной Африки составляли различные берберские племена. В результате арабского завоевания значительная их часть усвоила арабский язык. Берберские языки сохранили лишь отдельные группы населения: рифы, шлех — в Марокко, кабилы — в Алжире, туареги-кочевники Западной Сахары и среднего течения Нигера.
Доарабское население Северной Африки составляли различные берберские племена. В результате арабского завоевания значительная их часть усвоила арабский язык. Берберские языки сохранили лишь отдельные группы населения: рифы, шлех — в Марокко, кабилы — в Алжире, туареги-кочевники Западной Сахары и среднего течения Нигера.
Берберы отличаются от арабов угловатыми чертами лица, прямым по форме спинки носом, более широкой головой, менее высоким ростом. Однако такое разграничение не может быть строго проведено: как среди арабов, так и среди берберов проявляется большое разнообразие в отдельных признаках. В антропологической литературе неоднократно отмечалось наличие среди берберов светлоглазого и сравнительно светловолосого элемента. Наиболее светлы рифы. Вопрос о происхождении этого светло-пигментированного типа неясен. Его появление приписывали вторжению вандалов с севера. Однако следует заметить, что еще на древнеегипетских фресках встречаются изображения светлоглазых ливийцев.
Внимание антропологов давно привлекал вопрос о физическом типе древних обитателей Канарских островов — гуанчей. Выяснилось, что среди них отчетливо различаются два варианта. Один — узколицый, средиземноморский, другой — более широколицый, с более низкими орбитами, более отчетливыми надбровными дугами, в некоторой степени сходный с мезолитическим населением Северной Африки.
Население Мадагаскара неоднородно по антропологическому составу. Главные элементы, из которых оно сложилось, — южно-азиатский и негроидный. Первый наиболее выражен у мерина, второй — у бара, сакалавов и некоторых южных групп населения острова. В целом малагасийцы характеризуются преобладанием узкого разреза глаз, приподнятостью их наружного края, выступани-ем скул; эпикантус встречается редко; у большинства кур-
чавые волосы, уплощенный и довольно широкий нос, прогнатизм выражен в разной степени, цвет кожи очень варьирует. Если происхождение южноазиатской примеси бесспорно связывает малагасийцев с Индонезией, то вопрос о генезисе экваториального компонента долгое время оставался совершенно неясным.
Исследования М. Шамля (1958) приводят к выводу, что темнокожее население Мадагаскара происходит из Африки. Это подтверждается большим сходством бара и некоторых других малагасийцев не с меланезийцами, ас неграми Мозамбика и Южной Африки по краниологическим признакам, по соматологическому типу, по кожным узорам на концевых подушечках пальцев; кроме того, они оказались похожи на африканских негров по направлению ладонных линий.
Европа
В лингвистическом отношении большая часть населения Западной Европы относится к индоевропейской системе языков, которая в целом подразделяется на следующие группы: индийскую, иранскую, армянскую, славянскую, летто-литовскую (балтийскую), греческую, албанскую, романскую, кельтскую, германскую. Меньшую часть составляют финно-угорские языки. Баски, живущие в крайнем юго-восточном углу Франции и прилежащей области Северной-Испании, говорят на языке яфетической группы. Среди народов Балканского полуострова небольшими группами разбросаны турки-османы. Довольно значительный процент в некоторых странах Западной Европы составляют евреи, лишь условно относимые к семитам, так как они в большинстве уже не знают древнееврейского языка и говорят на языке той страны, где ныне обитают, или на особом диалекте одного из германских языков с сильной примесью слов из польского, древнееврейского и других языков. Другой народностью, рассеянной по разным странам Западной Европы, являются цыгане, наиболее многочисленные в Румынии и Венгрии.
Зак. 4ВВ
1) форма волос — широко волнистые или прямые, тон кие и длинные; 2) переносье высокое или выше среднего, горизонталь ное основание носа,… 3) ортогнатность лицевого черепа; .Температурные адаптации
Эффективность адаптации организма зависит от степени нарушения гомойотермии. Адаптивные температур-| ные приспособления человека бывают трех… 1. Общие физиологические приспособления, связанные с функцией… v 325Перегревание
| во-вторых, путем потоотделения. Роль системы кровообращения состоит в усилении тока крови через кожу, которое становится возможным благодаря… | кожных сосудов, а также увеличению минутного объема | сердца, сопровождающегося увеличением пульса. Тепло, поступающее в избытке к поверхности тела, рассеивается за счет…Охлаждение
v 331 ilL. зования, чтобы человек мог находиться на холоде несколько часов подряд. Теплоизолирующая способность одежды из…Адаптации к коротковолновому излучению
столкнулся с новой большой экологической опасностью, обусловленной освобождением и накоплением искусствен-ных радиоактивных веществ.Ультрафиолетовое излучение
Распространение интенсивности ультрафиолетового излучения на земном шаре в точности неизвестно. Уровень его в значительной мере зависит от… ч, ' 341 Под действием ультрафиолегетовых лучей кожный покров человека приобретает темнщый цвет. В основе загара лежит ряд…Ионизирующее излучение
Радиоактивные вещества попадают в организм человека через дыхательные пути и желудочно-кишечный тракт. Некоторые из них, например стронций-90 и… - Самые серьезные биологические последствия облучения связаны с мутагенным… <• 345Адаптации к высокогорью
Некоторые из этих функциональных сдвигов вызывают мысль об изменениях и в морфологических признаках высокогорных популяций; основанием этому… от их расовой и этнической принадлежности, характерно замедление ростовых… Пока еще очень мало обращено внимания на изучение морфофункциональных особенностей высокогорных групп взрослого…Зак. 486
Каждое сообщество вынуждено приспосабливаться к различным ввдам стресса, создаваемого самыми разнообразными факторами внешней среды.- Именно…Плотность населения и его численность
Каждая экономическая группа испытывает на себе действие многих различных факторов. Плотность популяции в данных условиях среды зависит от ряда… 1) от характера климата, почвы и рельефа; 2) от флоры и фауны;Регуляция численности популяций
Динамика численности человеческих популяций может быть весьма разнообразной: они могут увеличиваться или уменьшаться, а также оставаться стабильными. Уже в первобытных человеческих сообществах существовала
V 357
 | |||
 | |||
| B№OfM |
тенденция к сохр^юнюочислешюстн шзщлзщш на определенном уровне- Здесь следует упомянуть о системах естественной регуовщшь, действующих & Основные принципы этих систем^ еформулнр
s эко-
логами Элтоном, Нинольсоиами Соломоном, следующие

 на самом деле означает, что колебания численности
на самом деле означает, что колебания численности
2. Стабильный средняя уровень плотности имределяег-
ея емкостью системы (примером могут служить по-
нуяяцшг аборигенов Австралии).
3. Процессы, которые ограничивают диапазон колеба
ний к экосистеме постоянной емкости, — это внут
ренние процессы; их называют процессами,, завися
щими от плотности.
4. Внешние события могут резко нарушить равновесие
- к экосистеме, во они не участвуют в установлении
стабильности.
5. Люба»да1шая Э]тас1«л«ма имеет ««игн*вльнук)»чис-
населения (Элтон, 1941)» к процессы, регу-





 ошггамума (кторьш, однако, не всегда даствжаетеявг . данных условиях^.
ошггамума (кторьш, однако, не всегда даствжаетеявг . данных условиях^.
Эта принципы применимы в ж человеческим попу-лшрш»с, нос известными огравнлениямя. Вэ-нервыхт чне-
 лящшрегулнрустся нетольг-ко биологическими, но в культурными фанторамв. Действие последних может быть целенаправленным т во во многих сообществах оно настолько тесно переплетено с действием укоренившихся обычаев, что кажехса стань же безликим, как и проявление действия биологических факторов. Во-вторых, со временем система становятся все более способной к расширению. Е самый последний перв-од стало возможным непрерывное увеличение чвелешю-стипоиуляцйи без п|>нбляжения к равновесию.
лящшрегулнрустся нетольг-ко биологическими, но в культурными фанторамв. Действие последних может быть целенаправленным т во во многих сообществах оно настолько тесно переплетено с действием укоренившихся обычаев, что кажехса стань же безликим, как и проявление действия биологических факторов. Во-вторых, со временем система становятся все более способной к расширению. Е самый последний перв-од стало возможным непрерывное увеличение чвелешю-стипоиуляцйи без п|>нбляжения к равновесию.
Регуляторные процессы, зависящее от плотности, можйо раз делить на две группы—
В процессах первой группы ограничивающее действие изменяется с изменением численности оотгуляции — при возрастании численности оно усиливается, а при падении ослабевает. Так, нриувеличении плотности популяции усиливается борьба за основные источники нищи, воды, топлива "И сырья, за -освоение территорий н мест, пригодных для жилья. Увеличение плотности популяции может сопровождаться загрязнением окружающей среды или каким-либо иным ухудшением условий существования, что
б
д
распространению заболеваний как путем прямых к-онтак-
тов между людьми, так и вследствие загрязнения воздуха,
пищи, воды, домашней утвари. Ограничивающее действие
этих процессов ослабевает с уменьшением численности
популяции. ,.
Ограничивающее действие регуляторных процессов второй группы («инверсных») изменяется в направлении, противоположном изменению численности популяции. Это такие процессы, интенсивность которых с увеличением плотности популяций снижается и, наоборот, с уменьшением ее повышается. По мере увеличения численности популяция становится способной эффективнее использовать тпяфодяьюресурсь1, более успешно добывать себе средствах существованию, в особенности пищу. Появляются лучшие возможности для совместной шшъшрыбо-ловства, расчистки земель и проведения оросительных работ, строительства жилищ и т. д. Более многочисленное сообщество может лучше организовать взаимную защиту своих членов или обеспечить наступательные действия против других групп и животных. Следует учитывать также, что с увеличением численности популяции возрастает фонд генетической изменчивости. При уменьшении плотности все эти преимущества теряют силу: малочисленное, раз-


 ше шансов успешноконкурировать сдругими популяциями и менееспособно к проведению коллективных мероприятий.
ше шансов успешноконкурировать сдругими популяциями и менееспособно к проведению коллективных мероприятий.
В животном царстве эти две группы естественных ре-гуляторных процессов, действующих в противоположных направлениях, могут, по-видимому, поддерживать колебания численности популяции в узких пределах, совместимых с постоянным выживанием в данной экосистеме. По аналогии можно привести данные, свидетельствующие об относительной стабильности человеческих экосистем в неиндустриальных сообществах.
1. Археология предоставляет в наше распоряжение мно
гочисленные примеры того, как в определенных ус
ловиях обитания техника производства может оста
ваться неизменной в течение длительного периода
времени (Childe a., G. Clark). Анализируя медленное
изменение культуры каменного века, Чайлд справед
ливо утверждал: «До тех пор пока достигнутая сте
пень приспособленности к условиям среды остается
удовлетворительной, сообщество стремится быть кон
сервативным». Действительно, резкие изменения
жизни народа и его материальной культуры, устанав
ливаемые раскопками, обычно связаны с вторжени
ем новых популяций с иной культурой.
2. Этнография приводит данные, подтверждающие кон
серватизм тех современных сообществ, которые со
хранили многие примитивные черты (Forde). К при
меру, религиозные церемонии и традиции у австра
лийцев племени аранда подчиняются исключительно
строгой регламентации, и нарушители всегда сурово
наказываются. Экономическая деятельность таких
сообществ подчиняется строгим правилам и представ
ляет результат опыта, накопленного за долгое время
путем проб и ошибок. Строгое соблюдение границ
племенных территорий свидетельствует о стабильно-
сти популяции, так как ресурсы, необходимые для существования популяции, остаются неизменными втечение длительного времени.
3. Экономика примитивных сообществ почти полностью зависитот местныхресурсов. Стабильность популяции обусловлена негибким характером экосистемы. Для собирания плодов и охоты может потребоваться почти весь наличный запас энергии, который человек способен израсходовать в процессе работы, — примерно 6270—8370 кДж в день (эти цифры представляют расход энергии при непрерывном движении). Можно рассчитать, что плотность популяции в таких экосистемах должна составлять около одного человека на 3 км2. Переход к сельскому хозяйству приводит вначале к увеличению плотности популяции, однако значительные излишки при этом не накапливаются. Энергия, которую необходимо затратить на обработку 1,2 га земли в один год одним человеком, примерно равна энергии, затрачиваемой при использовании для охоты 200 га. По этому расчету плотность населения на стадии первобытного земледелия при равновесии может достичь 30 человек на 1 км2 или больше. 4. Мы располагаем весьма скудными сведениями о динамике численности населения в примитивных сообществах. Даже для относительно развитых сообществ, обладающих письменностью, часто отсутствуют соответствующие демографические данные. Наиболее надежными оказались данные о среднем числе дочерей, рождаемых одной матерью. Хорошим показателем может служить общая плодовитость (G. Clark, 1958). Последняя определяется числом детей, рождаемых в среднем одной женщиной за весь детородный период. При благоприятных условиях для воспроизведения потомства (т. е. когда каждая женщина выходит замуж в молодости, а вдовы имеют возможность вновь вступить в брак), если предположить
v 361
 к тому же, что климакс наступает после 20 лет супружеской жизни, общая плодовитость должна составлять примерно 8 детей. Это предполагает рождение одного ребенка за каждые 2,5 года замужества (учитывается возможность нескольких самопроизвольных абортов в период временной стерильности). При высокой смертности (когда лишь немногие переживают возраст 40 лет) общая плодовитость, измеряемая величиной 6—8, достаточна лишь для того, чтобы сохранить численность популяции на постоянном уровне. Известно, что в примитивных сообществах общая плодовитость часто бывает ниже 6 (табл. 9).
к тому же, что климакс наступает после 20 лет супружеской жизни, общая плодовитость должна составлять примерно 8 детей. Это предполагает рождение одного ребенка за каждые 2,5 года замужества (учитывается возможность нескольких самопроизвольных абортов в период временной стерильности). При высокой смертности (когда лишь немногие переживают возраст 40 лет) общая плодовитость, измеряемая величиной 6—8, достаточна лишь для того, чтобы сохранить численность популяции на постоянном уровне. Известно, что в примитивных сообществах общая плодовитость часто бывает ниже 6 (табл. 9).
Таблица 9
Общая плодовитость (число детей у матери) (Дж. Харрисон, 1979)
Собиратели и охотники-одиночки Австралийцы: западные области центральные области Индейцы Огненной Земли 4-6 5 …Процессы, регулирующие численность популяции
%- 363 Некоторые теоретики (Hawley, 1950) в своем анализе динамики популяции в… Размер популяции автоматически приспосабливается к изменениям природных ресурсов чаще всего за счет из- менения…Таблица 10
Меры по регуляции численности в различных популяциях (Дж. Харрисон, 1979)
Примечание: (+) — зарегистрированные случаи, (-) — не зарегистрированные случаи. время стали прибегать главным образом к механическим и химическим… Возможно, Карр-Саундерс преувеличивает необходимость общественных мер по ограничению рождаемости; ведь помимо высокой…Экологические показатели
личными критериями (в том числе такими, которые определяют уровень потребления, работоспособность, энергию или денежный доход, приходящийся на душу… Детская смертность. Наиболее важным показателем эффективности экологического… Средняя продолжягельность жизни. Этот показатель для человеческого общества каменного века и более ' позднего…Оо
Й а а я 8 8 й
2.8. Зубы........................................................ ............… Глава 3. Возрастная антропология................ ;..................... 50ЭТНИЧЕСКАЯ АНТРОПОЛОГИЯ
9.1. Расы животных и человека................................ 266 9.2. Раса и нация.............................................. „....... 270 … 9.3. Расообразование.................................................... 272– Конец работы –
Используемые теги: антропология0.041
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: АНТРОПОЛОГИЯ
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.156 сек.
Новости и инфо для студентов