рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биотехнологии
- /
- Основные положения хромосомной теории наследственности. Сцепление генов и кроссинговер. Генетические и цитологические доказательства кроссинговера. Генетические карты и принципы их построения.
Реферат Курсовая Конспект
Основные положения хромосомной теории наследственности. Сцепление генов и кроссинговер. Генетические и цитологические доказательства кроссинговера. Генетические карты и принципы их построения.
Основные положения хромосомной теории наследственности. Сцепление генов и кроссинговер. Генетические и цитологические доказательства кроссинговера. Генетические карты и принципы их построения. - раздел Биотехнологии, Основные Положения Хромосомной Теории Наследственности. Сцеплен...
Основные положения хромосомной теории наследственности. Сцепление генов и кроссинговер. Генетические и цитологические доказательства кроссинговера. Генетические карты и принципы их построения.
Хромосомы – ядерные структуры довольно плотной консистенции, интенсивно окрашивающиеся основными красителями. Хр.теор.наслед. - теория, согласно которой гены расположены в хромосомах в линейной последовательности и таким образом именно хромосомы представляют собой материальную основу наследственности, т.е. преемственности свойств организма в ряду поколений. Параллелизм в поведении генов и хромосом был отмечен еще Сэттоном и Бовери, он же послужил основанием для хромосомной гипотезы, а в дальнейшем – теории наследственности. В своих экспериментах они обнаружили параллелизм в поведении генов и хромосом при образовании гамет и оплодотворения. Эти наблюд. послужили основой для предполож., что гены расположены в хромосомах. Однако экспериментальное доказательство локализации конкретных генов в конкрет хромосомах было получ. Тольк. в1910 америк-м генети Морганом, кот. в последующ. г. (1911-26) обосновал хромос-ную теор. наслед-ти. Согласн этой теор., передача наследственной информации связана с хромосомами, в которых линейно, в опред-ной последоват-ти, локализ-ны гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности. Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установл-ы различия в наборе хром-м у организмов различ-х полов. При сопоставл. закономер-тей наследов-я признаков и поведения хр-сом в мейозе и митозе Морган сформулир-л следующ. основные положения:
носителями наследственности являются гены, находящиеся в хромосомах;
каждый ген имеет в хромосоме определенное место (локус гена);
каждая хромосома содержит десятки тысяч генов, кот-ые расположены в ней линейно;
гены в одной хромосоме называются сцепленными и образуют группу сцепления. Число групп сцепления равно гаплоидному набору хромосом;
гены в одной хромосоме наследуются совместно;
перекомбинация признаков происходит за счет независимого расхождения хромосом в мейозе и обмена му участками гомологических хромосом (кроссинговера);
мутации возникают за счёт изменения генов и хромосом.
Благодаря работам Моргана было достигнуто понимание явления «сцеплениегенов» - совместная передача двух или более генов от родителей потомкам. Объясняется тем, что эти гены лежат в одной хромосоме, то есть принадлежат одной группе сцепления и поэтому не могут случайно перекомбинироваться в мейозе, как это бывает при наследовании генов, лежащих в разных хромосомах. Число групп сцепления равно числу пар хромосом, т.е. гаплоидному набору хромосом. Закон сцепления: гены, находящиеся в одной хромосоме, образуют группу сцепления и наследуются по схеме моногибридного скрещивания. Разобрать это явление можно на следующем примере: если 2 гена полностью сцеплены, то дигибрид будет давать 2 сорта гамет- АВ и ав в отношении 1:1. Явление неполного сцепления генов (когда наряду с родительскими сочетаниями признаков в потомстве от скрещиваний обнаруживаются и новые их сочетания) было подробно исследовано Морганом и его сотрудниками и послужило обоснованием линейного расположения генов в хромосомах. Морган предположил, что сцепленные гены гомологичных хромосом, находящиеся у родителей в сочетаниях 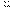 и
и  , в мейозе у гетерозиготной формы
, в мейозе у гетерозиготной формы 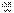 могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами
могут меняться местами, в результате чего наряду с гаметами АВ и ab образуются гаметы Ab и аВ. Подобные перекомбинации происходят благодаря разрывам гомологичных хромосом на участке между генами 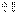 и последующему соединению разорванных концов в новом сочетании. Кроме сцепления генов в аутосомах, может происходить и сцепление признаков с полом. Так Морган скрещивал мух с белыми (w) и красными глазами (w+), результаты скрещивания зависили от того, кто из родителей имел белые или красные глаза – это реципрокные скрещивания. При скрещивании нормальной самки с белоглазым самцом все потомство было нормальным. Если же скрещивали белоглазых самок с красноглазыми самцами, то происходило расщепление 3 (кр):1 (бел). Т.о было выяснено, что красные глаза – доминантный признак, а белые – рецессивный. Результаты, полученные при скрещивании белоглазых и красноглазых мух, Морган объяснил предположением, что ген w находится в Х-хромосоме, тогда как У-хромосома генетически инертна. Такой тип наследования получил название наследования, сцепленного с полом. Т.о. ген w сцеплен с полом, т.е. находится в х-хромосоме. Гетерозиготные самки всегда красноглазы, что свидетельствует о рецессивности аллели w. В то же время самцы с аллелью w всегда белоглазы, в связи с инертностью У-хромосомы. Присутствие только одной аллели и в единичном числе у диплоидного организма называется гемизиготным состоянием, а организм – гемизиготой. Кроме того, Х-хромосомы закономерно переходят от 1го пола к другому – крис-кросс наследование.
и последующему соединению разорванных концов в новом сочетании. Кроме сцепления генов в аутосомах, может происходить и сцепление признаков с полом. Так Морган скрещивал мух с белыми (w) и красными глазами (w+), результаты скрещивания зависили от того, кто из родителей имел белые или красные глаза – это реципрокные скрещивания. При скрещивании нормальной самки с белоглазым самцом все потомство было нормальным. Если же скрещивали белоглазых самок с красноглазыми самцами, то происходило расщепление 3 (кр):1 (бел). Т.о было выяснено, что красные глаза – доминантный признак, а белые – рецессивный. Результаты, полученные при скрещивании белоглазых и красноглазых мух, Морган объяснил предположением, что ген w находится в Х-хромосоме, тогда как У-хромосома генетически инертна. Такой тип наследования получил название наследования, сцепленного с полом. Т.о. ген w сцеплен с полом, т.е. находится в х-хромосоме. Гетерозиготные самки всегда красноглазы, что свидетельствует о рецессивности аллели w. В то же время самцы с аллелью w всегда белоглазы, в связи с инертностью У-хромосомы. Присутствие только одной аллели и в единичном числе у диплоидного организма называется гемизиготным состоянием, а организм – гемизиготой. Кроме того, Х-хромосомы закономерно переходят от 1го пола к другому – крис-кросс наследование.
Морган выявил, что сцепление не всегда бывает абсолютным, сцепление может нарушаться. Причиной нарушения сцепления генов стал кроссинговер. Кроссинговер - взаимный обмен участками парных хромосом, происходящий в результате разрыва и соединения в новом порядке хроматид; приводит к перераспределению (рекомбинации) сцепленных генов. Т.о., кроссинговер — важнейший механизм, обеспечивающий комбинаторную изменчивость, а следовательно, — один из главных факторов эволюции. Кроссинговер, как правило, имеет место в профазе 1 мейоза, когда хромосомы представлены четырьмя нитями. Результат кроссинговера можно выявить по новому сочетанию сцепленных генов (если аллели гомологичных хромосом, участвовавших в К., были гетерозиготны). Частота кроссинговера му генами пропорциональна расстоянию му ними, т.е. чем дальше друг от друга расположены сцепленные гены, тем больше вероятность кроссинговера между ними. Биологическое значение кроссинговера: увеличивается комбинативная изменчивость, которая даёт материал для естественного отбора. С учётом процента кроссинговера составляются генетические карты хромосом. На таких картах нанесено относительное расстояние му генами, которое измеряется в морганидах. Одна морганида равна 1% кроссинговера. Генетическое доказательство кроссинговера. Генетически доказать кроссинговер можно на примере опытов Моргана по изучению сцепленного наследования. Проводилось скрещивание дрозофил с генами b (черное тело) и vg (зачаточные крылья). При скрещивании мух bbvgvg X b+b+vg+vg+, то в первом поколении все особи были нормальными дигетерозиготами. Далее проводилось 2 типа анализирующих скрещиваний: в первом скрещивали дигетерозиготного самца первого поколения с гомозиготной самкой. При этом происходит расщепление признаков в потомстве 1:1 (черные с зачаточными крыльями и серые с нормальными крыльями). В ходе этого было установлено, что кроссинговер у самцов не происходит. Во втором опыте брали для анализа гетерозиготных самок первого поколения (b+bvg+vg) и гомозиготных самцов (bbvgvg), то наблюдается иное расщепление - кроме родительских комбинаций признаков, появляются новые. Сцепление нарушается за счёт того, что гены в гомологичных хромосомах поменялись местами благодаря кроссинговеру. Этот опыт показал что у самок произошел кроссинговер. Т.о. можно говорить о том, что у самцов кроссинговер не происходит, а у самок – происходит. Гаметы с хромосомами потерпевшими кроссинговер, называют кроссоверными, а с непретерпевшими- некроссоверными.
Цитоло доказат. После того как генетич методами удалось установить явление кроссинговера, необход было получ прямое доказат-во обмена участками гомологич хр. Крейтону и Мак-Клинтон удалось получить у кукурузы форму, у кот гомологич хромосомы различ-сь мор-фологич-одна была нормальн,а другая несла утолщ на конце одного плеча, 2-е её плечо было удлинено.Эти особен-ти в строен пары хромосом легко обнаруж при цитологич исслед-ях.
Генетич карты сцепления представляют собой одномерные схемы взаимного располож ге-нетических маркеров на индивидуальных хр. Они составлены пока лишь для: дрозофилы, кукурузы,томатов,мыши,нейроспоры,кишечной палочки. Г.к. составляют для каждой пары гомологич хром-м, группы сцепления нумеруют. Для того, чтобы составить к. необходимо изучить закономерности наследования большого числа генов. При составлении г.к.указ-ся гр. сцепления, полн или сокращ название генов, расстояние в процентах от одного из концов хр-мы, принятого за нулевую точку. Создание г.к. позволяет предсказывать характер наслед признаков, гены кот нанесены на карту, а в селекционной работе облегчает подбор пар для скрещ. На практике именно г.к. дают возможность локализ сложные г-е маркеры на первых этапах исследования и дают возможность их дальнейш изуч. Единица расстояния между генами – морганида (1%кроссинговера).На первом этапе картирования опр-т принадлежность гена к гр. сцепления. Число группа сцепления равно гаплоидному набору хро-м. Крайнему гену, наиболее удаленному от центромеры, соответствует нулевая, исходная точка. Удаление других генов от 0-точки измеряется в морганидах. Построение генетических карт имеет большое значение, знание их для культурных растений и животных позволяет спланировать селекционный процесс, что способствует получению надежных результатов в короткие сроки.
2..Современные представления о гене. Доказательства генетической роли ДНК. Структура ДНК и механизмы репликации.
Ген-элементарная структурная и функциональная единица наследственности.Ген рассм-ся как отдельная сист в пределах молекулы ДНК и как часть целого генотипа.Откр-я 40-50хг доказали сложное стр-е гена.1)явл-е множественного аллелизма-гены,кроме 2 альтернативных сост(А=а),дом-го и рецес-го,могут иметь более чем 2 сос. 1928гСеребровский,Дубинин,Аголом откр.яв-ие ступенчатого аллелизма,результатом стала центральная теория гена-«ген дробим,представ.собой сложн.комплексное образ-е,содержащее элементар-е единицы,к-е расп-ны внутри гена в линейном порядке.Участки гена спос.самомт.муировать. Оливер,Льюис,Грин откр.яв-епсевдоаллелизма,к-й свидет-ет о слож.строен. гена.Бензер предлож.комплементационный критерий аллелизма-цистранс тест-позвол.устан-ть аллельность или неаллельность 2-х рецессив.мут-й,эти мутации у диплоид.орган-в нах.в 2-х сос1)цис-пол-е-в 1гомологе нах.оба мутационных гена.2)транс-полож-фенотип д.б. разным:норма и мутант.Т.о. ген имеет сложное строение,представляет собой единицу,далее делимую при кроссенговере,и не представляет собой единицы мутирования.
Генетическая роль ДНК
Прямые док-ва: в 1928 Гриффитс обнаружил явление трансформации. При совместном введении в мышей вирулентных пневмоккоков штамма S(с полисахаридной…Структура ДНК и мех-мы репликации
Репликация. Расплетение нитей ДНК происходит при воздействии белка-геликазы. Имеется особая единица репликации-репликон. У кишечной палочки вся…Генетика пола. Хромосомный и балансовый механизм определения пола у животных и человека. Генетика человека. Методы изучения генетики человека.
Признаки, по которым отлич-ся особи разных полов, делят на первичные (морф-ие и физио-ские особенности организма, которые обесп-ют образ-е гамет и… Хромосомная теория: Было высказано предложение, что Х- и У-хромосомы имеют… Балансовая теория: механизм определения пола, первоначально открытый у дрозофилы. Определяется по отношению числа…Закономерности наследования признаков при взаимодействии генов. Аллельные и неаллельные взаимодействия.
Выбор гороха посевного был продиктован следующими условиями: 1) много сортов, четко различающихся по ряду признаков; 2) растения легко… 1 закон Моногибридного скрещивание (родители отличаются по 1 признаку) при анализе F1 все гибриды одинаковы н-р по…Изменчивость. Типы изменчивости, их закономерности. Мутационная теория Де Фриза. Классификация мутаций на основе изменения генотипа и их краткая характеристика.
Изменчивость-свойство живого организма.Способность преобразовывать новые свойства и признаки.Различают:наследственную и ненаследственную изменчивость.Оценка разнообразия признаков-изменчивости у особей всегда производится по их фенотипическому проявлению.Однако причины их фенотипического разнообразия могут быть разные: различие генотипов или разнообразие условий среды,определивших варьирование в проявлении признаков у организмов одинаковых генотипов.По этому принципу выделяют два типа изменчивости: генетическая и фенотипическая.Г/и-изменение свойств и признаков организма м.б обусловленно изменением гена или других элементов генетического аппарата клетки.Такие изменения называют мутациями(пример: появление в потомстве гомозиготных белых кроликов черного,у остистой пшеницы безостых форм,у зеленой водоросли хлореллы салатных).Изменчивость м.б обусловленна и различной комбинацией генов,что может привести к появлению новых признаков или к новому их сочетанию.Это комбинативная изм-ть.Фенотип.изм-ть.В процессе индивидуального развития наблюд., закономерные изменения морфологических,физиологических,биохимических и др.,особенностей орг-а.Время и порядок появления этих изменений в онтогенезе строго определяются генотипом.Такую изм-ть назыв., возрастной или онтогенетической(закономерное и постепенное физическое и умственное развитие человека).Разнообразие в проявлении одинаковых генотипов в различных условиях среды-модификационная изм-ть.
Сист-ое изуч-е мутаций нач.с 20 в.Первую мутационную теорию изложил в 1901 г.,Гюго Де Фриз.Проводил скрещивание растений и обнаружил появление новых признаков,кот-х не было у рода этого раст-я.Мутация-скачкообразное,прерывистое изменения наследственного признака.Положения мут.теор-и Де Фриза:1.М-я-случайное скачкообразное изменение признака;2.М-я наследственные изменения.3.М-я качественное изменение признака не образ., непрерывного ряда изменчивости,вокруг среднего значения.4.М-я м.б полезными,вредными,нейтральными.5.М-я могут возникать повторно.
Не точности Де Фриза:у растен., обнаруж.,частое появление мут-и,а у др., раст нет.Мнение о том,что сущ-ют мут-е.периоды и периоды относительного покоя ошибочно.1920 г.,Вавилов-закон гомологичных рядов наследственной изменчивости.Он обнаруживает сходство признаков в пределе семейста злаков.З-н: роды и виды генетически близкие характеризуются сходными рядами насл.изм-ти,с такой правильностью,что зная ряд изменчивости в пределах одного и того же вида можно предположить появление тиких же мутаций.Мутационный процесс по способам возникновения: спонтанный(ест.условия,без вмешат-ва чел-а) и индуцированный(возн., в процессе эксперимента,в усл-ях контролируемых чел-м).
Различают:Генные мут-и,связанные с изменением строения отдельного гена(мут-я гена от дикого типа к новому состоянию-прямая,а от мутантного к дикому-обратная(реверсия гена)).Хромосомные мут-и:связ.с изм-ем стр-ры и функц. Отдельных хром.2вида:1внутрихром:делеции(потеря сред.уч-ов),дефишенсы(потеря конц.уч-в);дуплекации-линейные повторения отдел.уч-в генов;инверсии-разворот сред.уч-ка хром.2траслокация-обмен м/у негомологичнымихром. Геномные мут-и-св.с изм-м числа хром.. Полиплоиды –орг-мы,в кот-х произошло умножение гаплоидных наборов. Анеуплоиды-орг-ы у кот-х число хромасом не является кратным гаплоидному.Полиплоидия-геномная мутация,сост в увеличен.,числа хромосом,кратному гаплоидному.Различают:Автополиплоидию полиплоиды,возникающие на основе умножения геномов одного вида;аллополиплоидия- полиплоиды,возникающие на основе умножения геномов разных видов.
Мутационная теория Де Фриза
в 1901 году Г.де Фриз сформулировал мутационную теорию, к-я во многом совпала с теорией гетерогенеза русского ботаника Коржинского. Так что справедливо говорить о мутационной теории Коржинского - де Фриза. В соответствии с определением де Фриза мутации – явление скачкообразного прерывистого изменения наследственного признака.
Основные положения мутационной теории де Фриза:1. Мутации –случайные качественные изменения какого-либо наследственного признака . 2. Мутации устойчивы изменения, передаются в ряду поколений в таком виде в каком они возникли у данного организма. 3. Мутации м.б. полезными, вредными, нейтральными 4. одни и те же мутации в природных популяциях могут возникать многократно. 5. вероятность обнаружения мутаций зависит от числа исследованных особей 6. Мутации – качественные скачкообразные изменения и не обр-т ряд изменчивости вокруг среднего значения какого-либо конкретного признака. де Фриз создал свою мутационную теорию на основе экспериментов с растением энотера. Но он не получил мутаций, он наблюдал результат комбинативной изменчивости, т.к. формы с к-ми он работал оказалисьсложными гетерозиготами по транслокации.
Классификация мутаций на основе изменения генотипа и их краткая характеристика
Мутации – скачкообразные изменения наследственного признака.
Классификация мутаций по генотипу: генные, хромосомные, геномные и цитоплазматические.
Генные (точковые) мутации – мутации связанные с изменением строения и функций отдельного гена. Единицей измерения явл-ся отдельный ген. Ген А может мутировать в состояние а1, а2, аn . серия измененных состояний одного и того же гена - серия аллельных генов, а само явление называется множественным аллелизмом. Любая аллель такой серии может возникать мутационно, от аллели дикого типа или любого другого члена данной серии.
А-Т↔ Г-Ц
Т-А↔ Ц-Г а)транзиции-замены пар нуклеотидов к-е не изменяют ориентации: пурин-пиримидин в пределах пары, это приводит к появлению новых генов. Б)Трансверсия – замена пар нуклеотидов, изменяющие ориентацию. В) вставка лишней пары нуклетидов г)выпадение пары нуклеотидов
У человека известна серия аллелей: IA IB I0
Группа АВ соответствует генотипу IAI B ,А – IAI A , В – I B I B ,0 – I0I0
Явление множественного аллелизма увеличивает резерв мутационной изменчивости, имеет приспособительное значение в эволюции.
Цитоплазматические – изменения плазмогенов, приводящие к изменению признаков и свойств организма. Цитоплазматические мутации стабильны и передаются по наследству из поколения в поколение. Но обнаружение их затруднено, т.к. одноименные цитоплазматические структуры встречаются в клетке в большом количестве. Для проявления рецессивной мутации в фенотипе необходимо, чтобы все соответствующие цитоплазматические структуры в клетке были мутантными. Природа цитоплазматических мутаций м.б. различной. Есть 2 типа изменений: 1. утрата структур. Пример: мутантные штаммы эвглены, к-е пи длительном выращивании в темноте утрачивают пластиды (пластидная мутация). Она необратима и приводит к изменению ряда свойств эвглен. 2. мутанты, у к-х цитоплазматические структуры функционально изменены. Н-р: дыхательные мутации у дрожжей. Спец. исследованиями было доказано, что митохондрии мутантных дрожжей не содержат цитохромоксидазы. Мутантные плазмогены м.б. рецессивными или доминантными. Рецессивные плазмогены сохраняются и воспроизводятся в клетке, но их проявление подавляется доминантными. Цитоплазматические мутации к-е определяют фенотип возникают спонтанно, частота их возникновения не велика. У свеклы некоторые клетки имеют мутантные желтые, белые пластиды, т.е. лишены нормальной зеленой окраски. Наличие Ц.мутаций обогащает генофонд вида, служит доп.резервом изменчивости в эволюции.
Хромосомные - Мутации к-е связаны с изменением структуры, функций отдельной хромосомы. Единицей изменения яв-ся отдельная хромосома. Делятся на внутрихромосомные; межхромосомные.
Внутрихромосомные – дефишенси-потеря концевых участков хромосомы. Пример: тяжелое наследственное заболевание – синдром кошачьего крика, плач ребенка похож на мяуканье – это синдром 5-й хромосомы. делеции – потеря средних участков хромосом. АВСDEF→ АВСEF. Д потерялся. Дупликации-линейные повторы одинаковых участков в конкретной отдельной хромосоме. Дупликации могут происходить в пределах одной и той же хромосомы, или сопровождаться переносом копии участка генет-го материала на др. хромосому.(ABCDE →ABCBC ;ABCCB).инверсии – разворот среднего участка хромосомы на 180°.( ABCD →ACBD)инверсии проявляются в фенотипе в виде видимых простым глазом мутантных признаков с рецессивным эффектом, но многие инверсии могут не иметь видимых проявлений.
Межхромосомные:транслокации -обмен ген-го матер-ла между не гомологичными хромосомами.транспозиции – изменение местоположения гена внутри одной хромосомы.
Геномные мутации связаны с изменением числа хромосом. 2 типа геномных мутаций: анеуплоидия, полиплоидия(эуплоидия). Анеуплоидия –мутации связанные с изменением числа хромосом, не кратного гаплоидному. 2n+1-трисомики; 2n-1-моносомики.ээти мутации проявляются в виде хромосомных болезней. особенно у чел-ка и жив-х лишняя хромосома приводит летальности. Пыльцевые зерна. Содержащие лишнюю хромосому, не функционируют, а мегаспора и яйцеклетка жизнеспособны и с с лишней хромосомой. У диплоидных высших растений моносомия летальна. У дрожжей-сахаромицетов моносомики жизнеспособны.
Моносомия известна у жив-х, особи мух с одной хромосомой 4 жизнеспособны, но они имеют меньшие размеры, менее плодовиты, имеют ряд морфологических отклонений от дикого типа.
Полиплоидия.- связаны с изменением целого числа хромосом. 2n→3 n →4 n →5 n →..10 n.Полиплоидия широко распространена в растительном мире, реже в животном мире. П-важнейший мех-м видообразования. Различают сбалансированные полиплоиды с четным числом наборов хромосом: 4n, 6n,8n и несбалансированные полиплоиы: 3n, 5n,7n. Полиплоидные формы отл-ся от прародителей большей мощностью, имеют более крупные листья, цветки, семена. Полиплоидные клетки и их ядра также крупнее. Для многих растений известны полиплоидные ряды. Они включ-т от 2 до 10n. Различают 2 типа П: 1.автополиплоидия- умножение одного исходного генома АА →ААА →АААА. У полиплоидов особый мейоз(Aaaa x Aaaa- расщепление по фенотипу 35:1, по генотипу 1:8:18:8:1). Полиплоидизация повышает фенотипическую, генот-ю изменчивость в растительном орг-ме. 2.аллополиплоидия.- умножение хромосом(разных геномов). Аллополиплоиды возникают в рез-те межвидовой, межродовой, отделенной гибридизации.(ААВВ-аллотетраплоид, ААВВСС-аллогексаплоид).Пример Карпеченко создал искусственный плодовитый гибрид между капустой и редькой. У капусты, редьки 2n=18; →18В+18R. Обильно цвела , но семян не давала, т.к у этого растения обр-ся несбалансированные гаметы.
Генетика как наука. Основные понятия, законы и методы генетики. Менделизм.
Генетика-это наука кот-ая изучает наслед-сть и измен-сть всех живых существ начиная от вирусов до чел-ка. Генетика как наука яв-ся самой молодой среди всех биол-х наук.
Наследственность-св-во организмов передавать при размножении информацию о своих признаках и особенностях развития потомству.
Единица наследственности- ген. Он опред-ет отдельный элементарный признак. При изучении наследств-ти различают 2 понятия: собственно наследственность и наследование. В понятие наследственности входят св-ва генов детерминировать построение специфической белковой мол-лы, развитие признака план строения ор-ма. Наследственность- отражает закономерности процесса передачи наслед-ых св-в ор-ма от одного поколения к другому.
Изменчивость- зак-ся в изменении генов и их комбинировании в процессе индивидуального развития ор-ов. Определенная система взаимодействующих генов наз-ся генотипом, система признаков и св-в организма- фенотип. Фенотип кот. Проявление генотипа, а генотип- сов-сть наслед-х задатков, опред.развитие признаков.
Изучение наследственности и измен-ти ведется на разных уровнях организации живой материи: молекулярным, хромосомном, клеточном, организационном, популяционном. Это исследование осущ-ся несколькими методами:
1)Генеологическии метод: позволяет изучает закономерности передачи наследственных признаков от одного поколения. При помощи этого метода устанавливают, что множество наследств-х признаков контролируется только 1 геном. Этот метод применяют только тогда когда известен порядок какой-нибудь из родителей и несут какой-нибудь аномальный ген.
2)Гибридологический анализ: система скрещиваний в ряду поколений дающая возможность анализировать закономерности наследственности и наследования отдельных ор-в и приз-в организмов при половом размножении, а так же изменчивость генов и их комбинаторику.
3)Цитологический метод: позволяет изучать материальные основы наследственности. Этот метод служит для исследования «анатомии наслед-ти».
4)Цитогенетический метод: позволяет изучить хромосомы в кариотипе чел-ка. Применение этого метода помогает открыть различные хромосомные болезни.
5)Популяцио. статический метод: позволяет изучить закономерности расспрос-я наследств-х признаков различных популяций. При помощи этого метода изуч.наслед.болезни, мутации разл-е популяции.
6)Биохимический метод: оно изучает болезни обмена в-в. Различные мутации могут привести к разрыву биохим.р-ий. Н-р обмен фенот-ов, если этот обмен нар-ся у чел-ка набл-ся альбинизм- отсуствие пегментов.
Задачи генетики: изуч. механизмов измен-я гена, воспроизвед-е генов и хромосом в каждом клеточном делении. Для генет-го анализа расследованию опред-х приз-ов при половом размно-ии необходимо производить скрещивание 2-х особей разл-ми призна-ми наз-ют гибридным первого поколения.
Главное достижение Менеделя заключается в том, что он сформулировал и применил принципы гибридологического анализа для проверки гипотез.
Правила Менделя:
1) Скрещиваемые организмы д.б. 1 вида,
2) четко отличаться по отдельным признакам,
3)признаки д.б. константы, т.е. воспроизводиться из поколения в поколение
4) необходимы характеристика и количественный учет всех классов расщепления.
1 закон Моногибридного скрещивание (родители отличаются по 1 признаку) при анализе F1 все гибриды одинаковы н-р по окраске – закон единообразия гибридов 1 поколения/ правило доминирования.
2 закон Дигибридное скрещивание в F2 проявляются особи как с доминатными так и с рецессивным признаками в соотношении 3:1- закон расщепления генов/ независимого наследования признаков.
После переоткрытия зак Менделя все эти правила были многократно повторены и подтверждены. Но с увеличением числа изученных пар признаков у различных организмов было установлено, что зак Менделя не явл-ся абсолютными а выполн-ся при определенных усло-ях. Н-р изучение ночной красавицы (не полное доминирование).
7.Основы генетики популяций. Факторы генетической динамики популяций.
1. Популяционная генетика- это сплав различных дисциплин: мокулярно-популяционная генетика, эволюция, генетика количественных признаков.
Методы исследования в популяциционной генетике.
Элементарное эволюционное явление - длительно направленное изменение генофонда популяции. Пример элементарного эволюционного явления:индустриальный… В соответствии с законом Харди-Вайнберга при относительно постоянных условиях… Закон Харди-Вайнберга:После одного поколения случайных скрещиваний частота одного локуса с двумя аллелями или частота…– Конец работы –
Используемые теги: основные, положения, хромосомной, Теории, наследственности, Сцепление, генов, кроссинговер, Генетические, Цитологические, доказательства, кроссинговера, Генетические, карты, нципы, построения0.177
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Основные положения хромосомной теории наследственности. Сцепление генов и кроссинговер. Генетические и цитологические доказательства кроссинговера. Генетические карты и принципы их построения.
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.229 сек.
Новости и инфо для студентов